
ホーム > 業務紹介 > 遺伝資源の収集・保存・配布 > 林木遺伝資源連絡会 > 林木遺伝資源連絡会誌 > 林木遺伝資源連絡会誌【2011 No.4】
更新日:2017年8月29日
ここから本文です。
スギ遺伝子保存林の再造成技術
-SSRマーカーによる遺伝的多様性評価からのアプローチ-
| 高橋誠1)・渡邉敦史2)・宮本尚子3)・矢野慶介4)・岩泉正和5)・小野雅子2)・篠﨑夕子5) | ||
| 1)森林総合研究所林木育種センター九州育種場 | ||
| 2)森林総合研究所林木育種センター育種部 | ||
| 3)森林総合研究所林木育種センター遺伝資源部 | ||
| 4)森林総合研究所林木育種センター北海道育種場 | ||
| 5)森林総合研究所林木育種センター関西育種場 |
1.はじめに
林木にかぎらず、さまざまな生物種は、それぞれの種内に多様な遺伝変異を有しています。種内に遺伝変異があるおかげで、環境の変化による自然淘汰が働いた時に、生物は新たな環境に適応した個体を生み出していくことができ、種として存続していくことができます。また、栽培生物においても人間の改良目標に応じて、有用な変異を有した個体を選抜することが可能です。このように考えると、生物の適応あるいは改良の源である遺伝変異は重要な資源(遺伝資源)であることが分かります。
林木は個体サイズが長大で、世代時間が長く、野生性が高く、一般的に高い遺伝変異を保持しているため、その遺伝資源の保存には、保存対象樹種の本来の生息域(天然林)内で保存する「生息域内(in situ)保存」に加え、天然林以外で林分として保存する「生息域外(ex situ)保存」、施設内で冷蔵庫や冷凍庫を用いて保存する「生殖質(花粉,種子)保存」といった異なる保存方法が併用されます。これは、成体として生息域内保存する場合、大面積を必要とし、また野外での保存のため保存環境を人間の手で制御することがしばしば困難であるなど、制約が大きいためです。
遺伝子保存林は生息域外保存の一つで、優良な遺伝子プールである親林分(採種源林分)から採種・育苗した苗木により造成された林分(後継林分)です。伐期に達した遺伝子保存林は、その林分から採種・育苗して後継林分を再造成すれば、伐採することが可能で、遺伝資源の利用と保存の両立を指向した方法といえます。遺伝子保存林はスギ,ヒノキなど、30余樹種を対象に、全国238箇所、約950haが設定されています。この内、スギの遺伝子保存林は139箇所、約260haに及び、箇所数では全体の58.4%を占めます(平成19年度末現在)。
遺伝子保存林の再造成では、元来保有している遺伝変異を減少させることなく、遺伝的な変異幅を維持して後継林分に引き継ぐことが重要です。再造成の方法として、各遺伝子保存林の全個体をさし木やつぎ木で増殖する方法も考えられますが、発根率などの増殖効率を考慮すると、事業的に全個体を無性的に増殖するのは現実的ではありません。採種方法に配慮して実生により再造成することが最も効率的な手法であると考えられます。
本報告では、スギ遺伝子保存林について、SSR(マイクロサテライト)マーカーを用いて遺伝的多様性を評価し、親林分と後継林分の遺伝的多様性を比較して、後継林分で遺伝的多様性が減少しているか?、減少がみられた場合にはどのような要因によるものであるかを検討しました。また、採種母樹数が後継林分の遺伝的多様性に与える影響を明らかにし、後継林分造成時に望ましい採種母樹数を検討しました。
2.材料と方法
本研究では、会津森林管理署管内に設定されているスギ遺伝子保存林の親林分(以下,AG10)と、そこから採種された種子を用いて、同署管内に造成された遺伝子保存林2林分(以下,AG21,AG22)の合計3林分を調査しました。AG10はクマスギ系の優良林分(人工林)です。
AG10,AG21及びAG22において、それぞれ72個体、57個体、44個体から針葉を採取しDNAを抽出しました。4つのSSRマーカーについて遺伝子型を決定し、1遺伝子座当たりの対立遺伝子数(Na)、アレリック・リッチネス(日本語に訳せば「対立遺伝子の豊富度」を表す指標といえます; AR)、平均ヘテロ接合体率の期待値(HE)と観察値(HO)といった遺伝的多様性などを表す指標を算出しました。これらはいずれも遺伝的多様性のレベルを表す指標で、値が大きいほど遺伝的多様性が高いことを意味します。詳細については、高橋ら(2010)を参照下さい。
また,AG10で得られた遺伝子型データを基に、採種母樹数が後継林分の遺伝的多様性に与える影響をシミュレーションにより検討しました。採種母樹数が変化した時の後継林分の遺伝的多様性を平均へテロ接合体率の期待値(HE)と1遺伝子座当たりの有効な対立遺伝子数(Ne)で評価しました。シミュレーションの詳細については高橋ら(2011)を参照下さい。
3.結果と考察
AG10,AG21,AG22の3林分の遺伝的多様性の指標を表-1に示しました。AG10と比較した時、AG21とAG22のHE,HOに顕著な減少はみられませんが、NaとAR (50)では、値が顕著に低下しています。AG21とAG22のアレリック・リッチネスはAG10に比べて明らかに減少しています(1%水準で有意)。
| 集団名 | Na | HE | HO | AR (50) |
|---|---|---|---|---|
| AG10 | 23.8 (5.2) |
0.882 (0.046) |
0.792 (0.117) |
16.42 (3.26) |
| AG21 | 16.0 (3.9) |
0.878 (0.022) |
0.852 (0.067) |
12.91 (2.71) |
| AG22 | 18.8 (4.3) |
0.860 (0.024) |
0.825 (0.064) |
13.68 (2.66) |
括弧内の数値は標準偏差
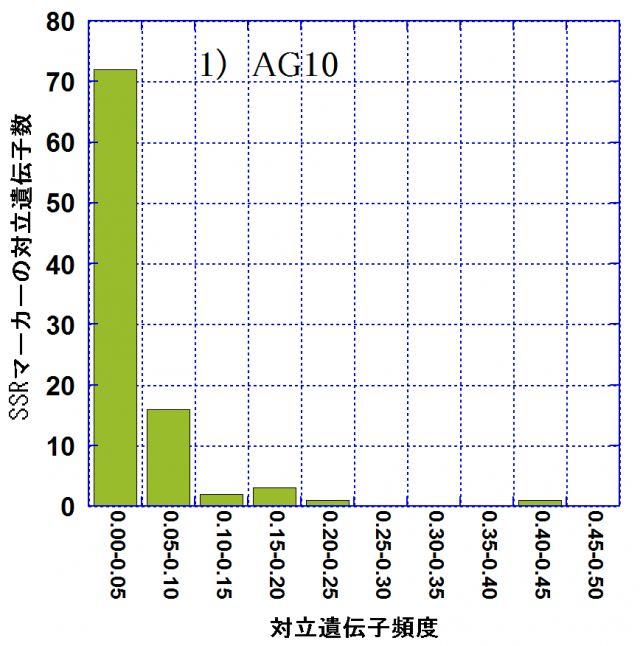
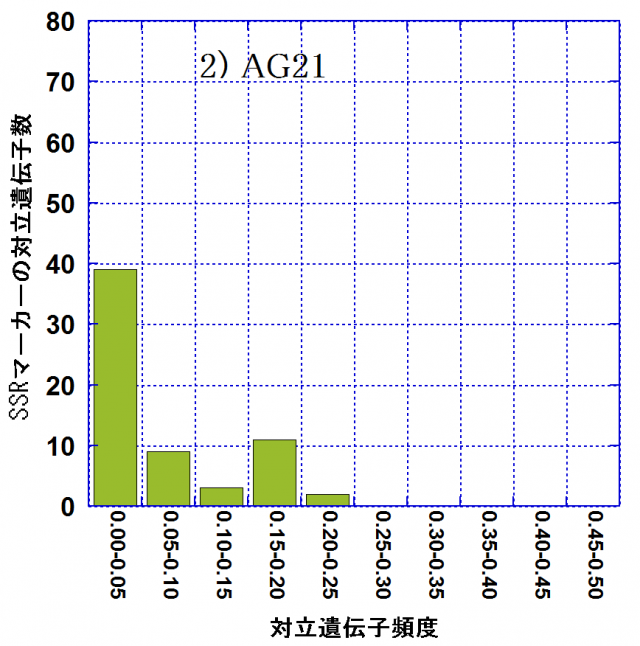
各林分での対立遺伝子頻度別の対立遺伝子数の頻度分布を図-1に示しました。頻度が0.05以下の対立遺伝子(アレル)をレア・アレルといいます。レア・アレル数は親林分(AG10)では72(75.8%)なのに対し、AG21では39(60.9%)、AG22では53(70.7%)でした。後継林分では親林分に比べてレア・アレルは数、割合ともに減少しており、アレリック・リッチネスの減少には、レア・アレルの消失が密接に関わっていました。その一方で、レア・アレル消失の影響を受けて、相対的に中程度の頻度(0.10~0.30)の対立遺伝子の割合はAG10における6.3%からAG21では25.0%、AG22では17.3%に増大していました。これらは弱度のビン首効果(個体数が急激に減少することにより遺伝的多様性が減少すること)の影響が現れたものと考えられます。AG21,AG22の造成には、AG10の27個体からの混合種子を用いて育苗した苗木を用いたとされています。後継林分造成の際の母樹数が、後継林分でのアレリック・リッチネスの減少に密接に関与している可能性が考えられます。
 |
 |
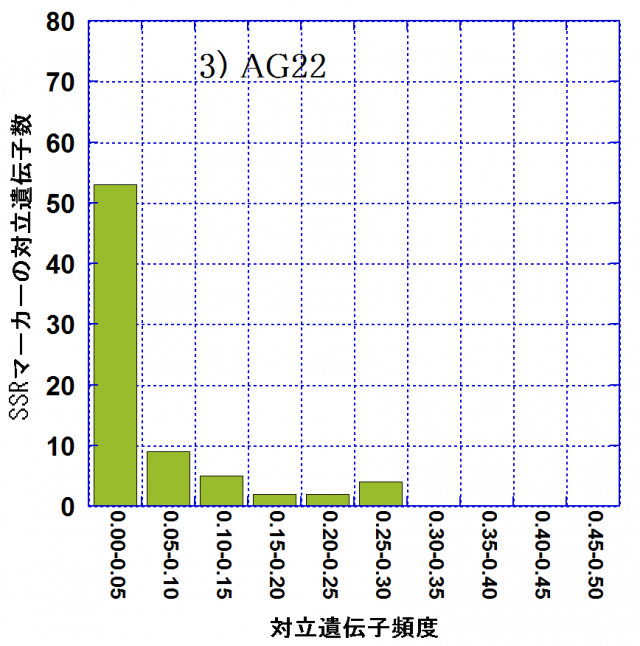 |
| 図-1 スギ遺伝子保存林の親林分(AG10)と後継林分2集団(AG21,AG22)における対立遺伝子頻度別の対立遺伝子数 | ||
次に後継林分の遺伝的多様性と採種母樹数の関係を明らかにするために行ったシミュレーションの結果について述べます。採種母樹数を10~100個体に変化させた時のヘテロ接合体率の期待値(HE)と有効な対立遺伝子数(Ne)の値を図-2に示しました。縦軸は採種源林分(AG10)が保有していた遺伝的多様性を100とした時のシミュレーションの結果(後継林分の遺伝的多様性の程度)で、ここでは遺伝的多様性維持率と呼ぶことにします。遺伝的多様性維持率の減少はNeで顕著で、特に採種母樹数が40個体以下で大幅な減少がみられました。
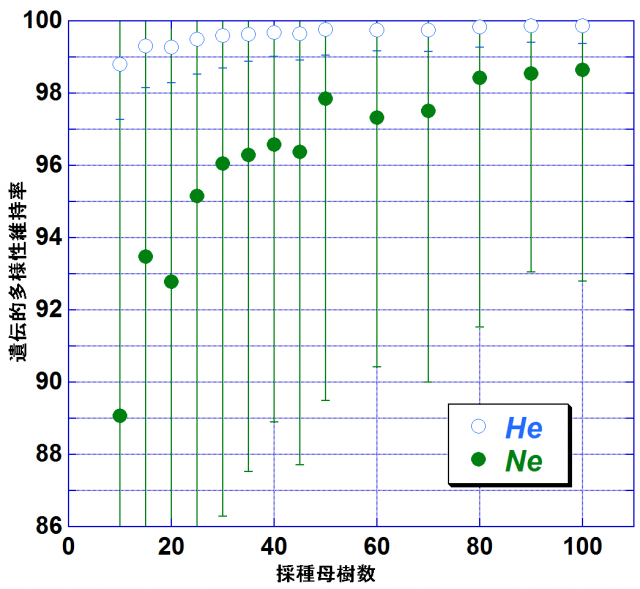
図-2 採種母樹数の変化に伴う1遺伝子座当たりの有効な対立遺伝子数(Ne)と 平均ヘテロ接合体率の期待値(HE)の遺伝的多様性維持率の推移
(エラーバーは1標準偏差を表す)
遺伝子保存林の再造成時に、元来保有していた優良なジーン・プールの遺伝的多様性を減少させることなく、後継林分に継承させることが望ましいわけですが、全ての遺伝的多様性を継承させることは困難なので、ここでは、元来の遺伝的多様性の95%以上を後代に継承させることを目標とした場合の望ましい採種母樹数について検討することにします。Neでみた場合、遺伝的多様性維持率は採種母樹数が25の時に約95%です。このことから採種母樹数は30個体以上にすることが望ましいと考えられます。また、遺伝的多様性維持率は、採種母樹数が50個体以上では顕著な変化は見られないこと、ならびに実際の採種作業にあたっては採種母樹数が増大するとともに必要な労力が増大することを考えると、採種母樹数は30~50個体にすることが望ましいと考えられます。
4.まとめ
遺伝子保存林の再造成時に後継林分に引き継がれる遺伝的多様性に与える採種方法、とりわけ採種母樹数の影響について検討しました。ここで得られた結果とこれまで知られている一般的な知見を勘案しつつ、下記に箇条書きしたような採種方法が望ましいと考えられます。
1) 採種は豊作年に計画する。必要に応じて森林管理署の許可を得てジベレリン処理を行う。
2) 採種母樹数は30~50個体とする。
3) 採種は高枝鋏などの用具を用いて各母樹から球果を採取する。しかし、それが困難な場合には、a. 森林管理署の許可を得て採種木を伐倒し球果を採種する、b. シード・トラップを用いて採種するなど、別の方策をとる。
4) 個々の採種母樹は林分中央付近に位置する個体の中から形質が優良な優勢木を選定する。各採種母樹は20~30mの間隔を空けることが望ましい。シード・トラップを使用する場合にあっては、シード・トラップの配置にあたり左記のような配慮をする。
5.謝辞
本研究の実施に当たっては、会津森林管理署の方々にご協力いただきました。ここに感謝の意を表します。
引用文献
1)高橋誠・渡邉敦史・宮本尚子・津村義彦・矢野慶介・岩泉正和・小野雅子(2010)スギ遺伝子保存林の遺伝的多様性の評価―福島県会津地方に設定されている採種源林分と子林分のSSRR―マーカーを用いた評価-.森林総合研究所林木育種センター平成21年版年報 83-85.
2) 高橋誠・渡邉敦史・宮本尚子・矢野慶介・岩泉正和(2011)SSRマーカーを用いたスギ遺伝子保存林の遺伝的多様性の評価―採種母樹数が遺伝子保存林分遺伝的多様性に及ぼす影響―.森林総合研究所林木育種センター平成22年版年報 92-95.
お問い合わせ
Copyright © Forest Research and Management Organization. All rights reserved.