
ここから本文です。
研究情報 No.39 (Feb. 1996)
巻頭言
森林研究の中の野生鳥獣
鳥獣研究室 北原 英治
野生鳥獣による深刻な森林被害や、林業と野生鳥獣との共存が久しく叫ばれています。特に、人工林化の進んだ森林を多く有し、しかもそれらの多くが人口密集地と接している西日本各地においては、被害防止策の早急な策定と野生動物との共存の声が大きいようです。しかし、これらの議論の行き着くところは「野生鳥獣の保護か人間の生活か」との論理になってしまい、「害獣は駆除して、益獣は増やそう」ということになります。これは判断基準を経済性にだけおいて論議するためです。最近の環境保護・動物保護に対する機運の高まりを考えますと、森林・林業の中での野生鳥獣についても、経済性と少し離れた観点での議論が必要と思われます。

写真-1 紀伊半島に隔絶して生息するヤチネズミ
森林の公益的機能の一つとして、山くずれや洪水を防止し、水源を涵養するという「森林の水土保全機能」が一般的に知られていますが、この外に、森林が野生鳥獣に対して、生息、繁殖、採食および休息などのいわゆる生活の場を提供する「野生鳥獣の保護・維持機能」をも評価すべきではないでしょうか。右に示したヤチネズミは、本州中部以北群と地理的に隔離して紀伊半島に分布していて、この地域群は日本列島の温暖化により寒冷樹林帯が北方へ退行したことによって、この地に取り残された残存個体群(レリック)と考えられています(写真-1)。このため、紀伊半島のヤチネズミは、日本列島の南限である大台が原に残るトウヒに並び、希少な動物と言えます。

写真-2 西日本に広く生息するニホンジカ
一方、農林業地に深刻な被害を引き起こしているニホンジカについて見れば、温暖寡雪で比較的下層植生の発達し、好適な餌環境の整った地域が広がっている西日本で急激に個体数を増やしたと考えられます(写真-2)。さらに、最近ではシカによるスギ・ヒノキ中・大径木の樹幹剥皮も見られ始めています。確かに、シカは極めて多様な植物を餌としますが、中・大径木の樹幹剥皮現象は、シカの餌環境が劣悪になりつつあることを示唆しています。今後、ネズミやシカなどの個体群管理もさることながら、野生鳥獣の生息を受け入れた森林の管理技術が必要と思われます。
研究紹介
森林植物の新しい親子判定法
造林研究室 井鷺 裕司
森林の中で生育する稚樹は一体どこからきたのでしょうか。親の樹木はどれだけ遠くまで、どれだけたくさんの種子をとばし、子孫を残すことができるのでしょうか。これは、森林の更新や保続的な管理という面で重要な問いかけです。また、近年人間の活動により多くの森林生態系が分断され、島状の分布様式をとるようになってきましたが、この様な分断化によって、どの様な種が更新不可能となって姿を消してゆくのでしょうか。また、残された種の遺伝的多様性に関して、どの様な影響がみられるのでしょうか。
これらの疑問に答えるために、いろいろな研究がなされてきました。例えば、樹木の種子生産量や散布量を調べるために、多数のシードトラップを林床に設置したり、ネズミやリスによる種子散布や貯蔵量を調べたりする事で、多くのことがわかってきました。しかし、これらの方法では種子を生産した雌としての樹木の役割を測定できても、遺伝的には同様に重要である花粉親、すなわち雄としての樹木の役割を明らかにすることはできません。また、種子が稚樹となり、どのように生存するかという点については別の調査が必要です。
現在生存している、様々な生育段階の樹木の父や母は一体どの個体なのか、直接的に明らかにできる手法はないのでしょうか。実はあるのです。
最近、犯人の識別や真の親子鑑定のために、遺伝子を担う物質であるDNAを、従来の犯罪捜査における指紋のように用いるDNAフィンガープリンティング法という手法が取り入れられているのをしばしば見聞します。 DNAフィンガープリンティング法には、いくつかの手法があります。初期のものについては手法やデータの解釈に批判もありましたが、手法の改良や解釈方法の改善に伴って、法廷でも信頼に耐えうる手法となってきました。
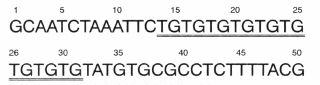
図-1マイクロサテライト部位の例
この図の場合、14番目の塩基から31番目の塩基までがマイクロサテライト部位で、TGというモチーフが9回繰り返されている。
私たちは、森林の樹木の更新過程を明らかにするために、DNAのマイクロサテライトと呼ばれる部位を分析しています。真核生物のDNAは、4つの塩基の配列が1細胞あたり数億から数百億並んでいて、これが様々な遺伝情報を伝えています。この中には、1~6塩基からなる短いモチーフが何度も繰り返されているマイクロサテライトと呼ばれる奇妙な部分があります(図-1)。マイクロサテライト部位は蛋白質のアミノ酸配列を決定する情報を持っていないため、この部分の繰り返し数は遺伝的に中立で、繰り返し数に変異が大きいことが知られています。図-1のように、あるマイクロサテライト部位前後のDNAの塩基配列がわかれば、この部分だけをPCRという手法で増幅し、マイクロサテライトの繰り返し数の違いを、個体間で比較することが簡単にできます(図-2)。
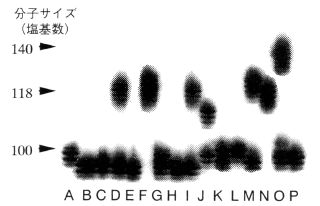
図-2マイクロサテライト部位の繰返し数の多型
16個体のシラカシ(A~P)についてマイクロサテライト部位を増幅し、サイズの違いを電気泳動で調べた。
この解析法は、親子判定を行う際に、他のDNAフィンガープリンティング法やアイソザイムを用いた分析法と比べて、非常に優れた特徴があります。まずは図-2で明らかなように、結果の像が非常にシンプルで解析がたやすいことです。図-2で各レーンは異なった個体に由来していますが、それぞれのレーンで、バンドは1本(A, B, C, E, F, G, H, K, L, N, P)もしくは2本(D, I, J, O)しかありません。1本の場合は両親から同じ長さのマイクロサテライト部位をもらった場合であり、 2本の個体は異なった長さのものをもらっています。また、マイクロサテライト部位は1細胞のDNA中に無数にあり、それぞれの部位で繰り返し数に変化が多いことも優れた特徴といえます。一つ一つの分析はシンプルでも、それを組み合わせる事で、非常に多くの情報を得ることができるのです。
この手法を用いれば、森林を構成する個体が一体どの個体から花粉をもらい、どの個体の種子に由来するものなのかを、直接的に明らかにできます。森林の動態を明らかにする上で非常に強力な道具となるでしょう。また、希少種の保全や人為撹乱下での種の保全に関しても、役立つ技術となるでしょう。
寄生バチは隠れた寄主をどのようにして発見するか?
昆虫研究室 浦野 忠久
寄生バチとは、さまざまな種類の昆虫に卵を産みつけ、それを子の餌(寄主)にして殺してしまうハチのことで、古くから作物害虫の天敵として用いられてきました。どの種類のハチも、人間にはとても見つけることのできないような場所にいる寄主を、実にうまく探し出すことができます。これには、寄主やその餌から発生する信号が関わっているのです。この信号にはさまざまなものがありますが、樹幹を食べる虫とそれに寄生するハチの例について紹介します。
枯れて間もないマツやスギなどの樹幹の中には、いろいろな昆虫が生息しています。とくに、樹皮下の形成層の部分が栄養価が高く、カミキリ、ゾウムシ、キクイムシなどの幼虫が数多く見られます。このような虫を樹皮下穿孔虫と呼び、もちろんこれらにも天敵である寄生バチが存在します。樹皮下穿孔虫の寄生バチの多くは長い産卵管を持ち、樹皮の外側からこれを差し込んで、寄主に麻酔液を注入した後に産卵を行います。しかし、ハチには寄主の姿を見ることはできません。では、ハチはどうやって樹皮下の寄主を発見するのでしょうか。
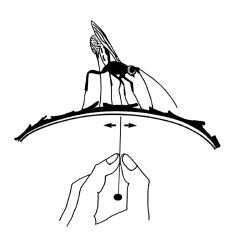
図-1DeLeon (1935) の実験
樹幹上のハチは、触角で樹皮の表面をたたく(あるいはこする)ようにしながら、寄主探索をします。このとき、樹皮下から伝わる何らかの刺激が、寄主の存在を知らせていることはまず間違いありません。これがどのような刺激であるかについて、これまでにいくつかの説が出されました。 1935年にDeLeon は、キクイムシの寄生バチに関して、寄主幼虫が樹皮下をかじるときの振動によって寄主を発見しているのではないかと考え、図-1のような実験を行いました。これは、丸太から剥いだ樹皮の上にハチをのせて、その裏側をピンの先でこするというものですが、ハチはこれによって産卵管を樹皮に突き立てる行動をした、と報告されています。
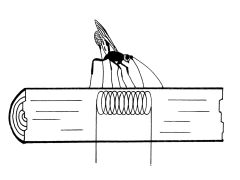
図-2Richerson and Borden (1972) の実験
1972年にRicherson and Bordenは、実際にハチが産卵行動を行った位置(すなわち寄主の真上)における樹幹表面温度が、その周囲より高いことを明らかにしました。そして、寄主に由来する代謝熱が樹幹表面まで伝わることによって、ハチは寄主の存在を知るのではないか、と考えました。これを確かめるために、丸太に電熱線コイルを埋め込んでこれを寄主に見立て、表面の温度を上げたところ、ハチはその真上で産卵行動を行った(図-2)、とされています。
その後Mills et al. (1991)は、寄主幼虫を取り出した丸太の上でも、ハチが産卵行動を起こすことや、新しい丸太に寄主を接種した場合、寄主が樹皮下を食い進むとともにハチの産卵行動が見られるようになることから、寄主の排泄した糞などの物質からの化学的な刺激が、寄主発見の手がかりである可能性が高い、と報告しています。
筆者も樹皮下穿孔虫の寄生バチを飼育していますが、その寄主探索行動を観察していると、樹皮の亀裂や、寄主が樹皮に開けた穴に対して、とくに念入りに触角をこすりつけているようです。寄主の存在を示す手がかりは、このような場所から伝わりやすいのかもしれません。では、それはどのような刺激なのか、上に挙げた3つの内のいずれか、それともまた別なものなのか、これから調査したいと思います。
連載
土の中の万華鏡(3)
植物が鉱物をつくる
土壌研究室 鳥居 厚志
植物が鉱物をつくる‥‥‥何のことでしょう?
ススキやタケ・ササ、シバなどのイネ科の植物は、土から珪酸を多く吸収するという特徴を持っています。これは他の種属の植物にはあまり見られない特色です。
水に溶け、根から吸収された珪酸は植物体の中を移動し、最終的に葉の表皮の細胞に沈着します。沈着した珪酸は、落葉して葉が分解した後も、細胞のかたちそのままに残り、土の中に蓄積されていきます。これが「植物珪酸体」と呼ばれるもので、植物がつくる一種の鉱物と言えます。
植物珪酸体はプラントオパールとも呼ばれ、宝石のオパールとほぼ同じような化学成分です。写真のように様々な形をしていますが、植物の種類毎に、ある程度形態の特徴があります。そこで、土の中の植物珪酸体を調べ、昔の植生を推定するのに利用されたりしています。
(写真は、本所森林環境部地質研究室 河室公康氏の提供による。粒径は10μm くらいから100μmくらいのものまである。)
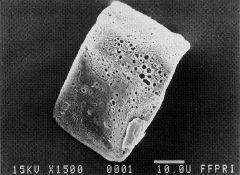
写真-1ススキ起源の「箱型」珪酸体
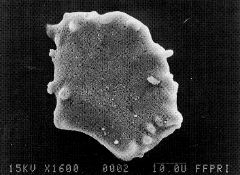
写真-2ササ起源の「ファン(扇)型」珪酸体
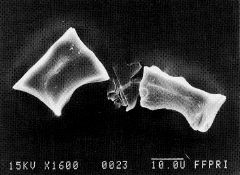
写真-3ササ起源の「タケ型」珪酸体

写真-4「棒状」珪酸体、起源不明
おしらせ
山城試験地に気象測定装置設置
森林気象の測定を行う装置が山城試験地内に2月中に設置されることになりました。これは、流域の谷地形における、特に風の動きを中心とした気象環境の分布を把握するために設けられるもので、日射計・純放射計・三次元超音波風向風速計・温湿度計等から構成されます。この装置をもちいて、林冠の形状や地形の効果によって、風の動きがどのようになるのか、などといった研究がすすめられます。
<WWWホームページ公開について>
37号でお知らせしましたWWWホームページ公開の件ですが、諸般の事情により遅れております。申し訳ございませんが、今しばらくお待ちください。
<訂正とおわび>
前号におきまして次の誤りがありました。おわびして訂正いたします。
4ページ下段:大阪営林署 → 大阪営林局
(注 … HTML化にあたり、訂正済みです)
Copyright © Forest Research and Management Organization. All rights reserved.