
ここから本文です。
研究情報 No.51 (Feb. 1999)
巻頭言
景観は管理すべきか?
-景観生態学的な視点から-
風致林管理研究室 大住克博
人が居住する景観、たとえば里山、田園などは、人と人を取り巻く自然環境の間の長い相互作用・共同作業の中で形作られてきたものです。例えば里山を代表するコナラ林は、単なる照葉樹林の二次植生ではなく、氷河期に南下していた植生が、人が森林を利用し撹乱し続けることで現在まで維持されてきたものであるといわれています。人は、薪炭や緑肥の採取の場としてコナラ林を生活体系の中に位置づけ、里山景観を作り上げてきました。このように景観には、森林の構造が人の活動により規定され、また集落や人の生業も森林のありようによって規定されるという、入れ子の構造が潜んでいるのかもしれません。
里山はそうだとしても、奥山の天然林の景観は人の活動と無関係ではないか? という疑問が、一方であるかもしれません。しかし、日本のように隅々まで人の居住する地域では、天然林と言えども、人との関わりの中で存在を考えるべきだと思います。なぜなら、間接的には下流の人口に左右される薪炭用、建築用の木材需要により、上流の天然林の運命が決まってきたからであり、また直接的には、天然林の多くは、藩政時代の土地所有、土地利用の線引きと、管理によって「残されてきた」ものだからです。これは木曽ヒノキ林などの美林ばかりではなく、東北地方の普通のブナ林についてもある程度当てはまることで、現在残存する天然林の分布が、小流域により偏在することなどが、それを物語っています。我々は「残されてきた」ということの中に、先人の意志も読み取るべきではないでしょうか。
さて、最近の森林景観に関わる議論の中には、何もしないことが肝要という意見もあるようです。景観の管理は、多様であるべきなので、その中には何もしないという選択肢もあると思います。しかし、それを放置の是認ととるのは短慮であると指摘しておかねばなりません。景観を維持してきた伝統的な土地利用や社会構造が失われてしまった中で、我々が今、自然との関係、その結果としての景観について迫られているのは、契約の破棄ではなく更改であると思います。放置だけでは、人と環境の共同作業である景観を明日に伝えられません。さらに、今世紀に入り、人間の活動が、環境を圧倒するほど肥大してしまっているという現状にも留意すべきでしょう。
それでも自由放任すべきという意見はあるでしょう。景観は人と自然の相互作用により巧まずして自己形成されてきたのだから、管理するべきではないと。景観管理が、往々にして押し付けがましい緑地や水辺の修景工事、公園造成に矮小化されてしまう現状を見れば、そのような批判も理解できます。しかし景観は自己形成されると言っても、その初期値や外部条件の設定が重要であり、それにより全く異なったパターンが生み出されることも考えられます。さらにその過程は、長い時間と膨大な作業を要するものであり、簡単にやり直しは効きません。
ここに我々が、景観に関わる諸関係を理解し、より賢い選択をしながら管理をしていくべき必要と責任があると思います。「何もしない」という選択も、この過程を経てなされるべきものでしょう。もちろん我々はここで、「景観はどうすれば管理できるのか?」という、次の重い問いに直面することになるのですが。
研究紹介
ホオノキのマイクロサテライト遺伝マーカー
造林研究室 井鷺裕司
1. はじめに
ホオノキは日本ではごく普通にみられる落葉性高木です。しかしながら群落内で優占する事は少なく、多くの繁殖個体密度は1ヘクタール数本程度で安定して維持されています。森林を構成する樹木にはブナやアカマツのように、群落内で林冠の大部分を占めるようなものがある反面、ホオノキをはじめ、トチノキ、カツラ、ヤマザクラのように比較的低頻度で安定して個体群が維持されている樹種も多くあり、森林の多様性の維持に貢献していると考えられます。このような樹種は、どのような範囲にわたって花粉を交換し、種子を散布しているのでしょうか?風媒、虫媒といったポリネーターの違いは受粉様式にどのような影響を持つのでしょうか?
2. マイクロサテライト遺伝マーカー
直接的には目で見えない花粉の動きや、完全に追うことはできない種子の散布範囲を明らかにするには何らかの遺伝マーカーに基づく情報を活用するのが理想的です。多くの遺伝マーカーの中でもマイクロサテライト部位の多型を利用した、いわゆるマイクロサテライト遺伝マーカーは、(1)共優性、(2)シンプルな解析、(3)多数の遺伝子座、(4)高度な多型、(5)PCR法による微量サンプルの解析、等の特性を持つため、上記のような解析には理想的なマーカーです。しかしながら、マイクロサテライト遺伝マーカーは、実際のサンプルの解析に進む前に、対象樹種ごとに、DNAの抽出と制限酵素消化、形質転換とライブラリー作成、スクリーニング、DNAシーケンシング、PCRプライマーのデザイン、等の作業を行う必要があり、この点は他のアロザイムやAFLP等のマーカーに比べると欠点であるといえます。
私たちはホオノキを対象に11遺伝子座のマイクロサテライト遺伝マーカーの開発を行いました(表-1)。いずれの遺伝子座も多くの対立遺伝子と高いヘテロ接合度を示し、個体識別を行うための優れたマーカーとして利用できることが明らかになりました。また、表-1に示したマイクロサテライト遺伝マーカーはホオノキだけでなく、同じホオノキ属の種、例えばタムシバ、コブシ、シデコブシ、オオヤマレンゲ等、日本産の落葉性の種でも利用可能でした。つまり、これらのマーカーを開発するのにはコストと手間がかかりますが、近縁種に関してはそのまま流用が可能なのです。
| 遺伝子座 | 繰り返し構造* | 対立遺伝子数** | ヘテロ接合度** |
| M6D1 | (CT)43 | 21 | 0.93 |
| M6D3 | (CT)22 | 22 | 0.94 |
| M6D4 | (CA)2(GA)15 | 26 | 0.90 |
| M6D8 | (CT)3C(CT)10 | 11 | 0.79 |
| M6D10 | (GA)14 | 6 | 0.73 |
| M10D3 | (GA)35 | 16 | 0.88 |
| M10D6 | (CT)11 | 7 | 0.81 |
| M10D8 | (GAA)6(GA)26 | 12 | 0.86 |
| M15D15 | (GA)16 | 5 | 0.69 |
| M17D3 | (CT)19 | 9 | 0.86 |
| M17D5 | (GA)19 | 10 | 0.86 |
| *クローンニングとシーケンシングを行った配列内での構造 **30個体の野生集団を解析して得られた値 |
|||
3. ホオノキ野生集団におけるgene flow
これらのマーカーを用いて野生集団において親子判定を行い、花粉や種子の散布範囲、gene flowの大きさ等について解析を行ってきました(図-1)。ホオノキは甲虫が花粉を媒介しており、花粉の移動はあまりないのではないかと考えられていましたが、予想に反して活発に移動していることが明らかになりました。林床の稚樹の遺伝子の半分が集水域外から来ていました。これらの実態は従来の手法では全く見えなかった事です。マーカー開発とフィールドワークの連携を保ちつつ作業を進めればマイクロサテライト遺伝マーカーは野外の樹木集団の更新過程を解明するに際してきわめて強力なツールとして利用できるでしょう。
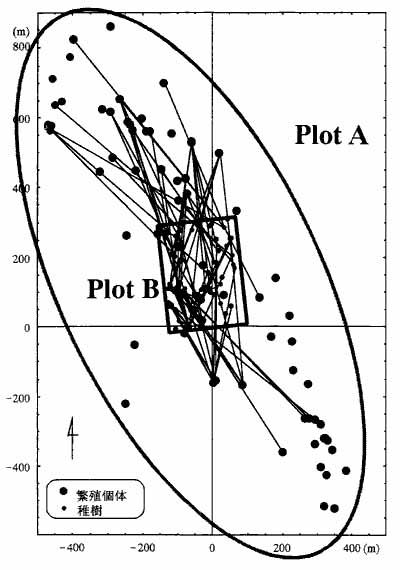
図-1 ホオノキ野生個体群の親子関係
北茨城市小川学術参考林内の1集水域をカバーするPlot A(面積69 ha)内のすべての繁殖個体とPlot B 内の稚樹の親子関係を線で結んだ。
雌雄を産み分ける寄生バチ
昆虫研究室 浦野忠久
前号(No.50)で同研究室の上田から、「半数倍数性」のキクイムシについての研究紹介がありました。今回は同じく半数倍数性で、雌雄を産み分ける性質を持った寄生バチについて紹介します。
寄生バチとは母親が餌となる昆虫(「寄主」と呼びます。)に卵を産み付け、孵化した幼虫が寄主の体内から栄養を摂取して、最終的には寄主を殺してしまうというもので、この生態を利用して害虫防除に役立つこともあります。ここで紹介するのはキタコマユバチという種類で、寄主となるのは枯れたマツの樹幹内(樹皮下)にすむカミキリムシ、ゾウムシといった甲虫類の幼虫です。雌バチは樹幹上で触角を用いて樹皮下の寄主を探り当て、腹部の先にある体長と同じくらいの産卵管を、樹皮の亀裂などを通して樹皮下に差し込みます。そして寄主に毒液を注入して動けなくした上で、卵を産み付けます。産む卵の数は寄主1個体につき1個です(これを「単寄生」といいます。)。孵化幼虫は寄主の体表面にくっついた状態で、寄主の内部組織を吸収して成長します。そして3日~1週間後には寄主の体を全て食べ尽くし、繭を作って蛹になります。
さて、単寄生バチ類では一般に寄主のサイズに依存した性比調節が行われることが古くから知られています。言い換えると、母親はサイズの比較的大きい寄主には雌の卵を産み、小さい寄主には雄の卵を産む、ということになります。これは寄生バチが半数倍数性で、雌雄の産み分けをする性質を持つことにより可能となっています。キタコマユバチにおいてもこのような性比調節が行われているかを実験してみました(図-1)。寄主を大小2つのサイズに分けて、それぞれのサイズを2個体同時に雌バチに与えて寄生させ、羽化した成虫が雌雄どちらになるかを調べました。その結果、サイズの小さい寄主に寄生した場合は、羽化成虫の性比が強く雄に偏ったにの対し、大きな寄主では雌雄ほぼ1:1となりました。この実験から、キタコマユバチが寄主サイズに応じて雌雄を産み分けている可能性が高いことがわかりました。
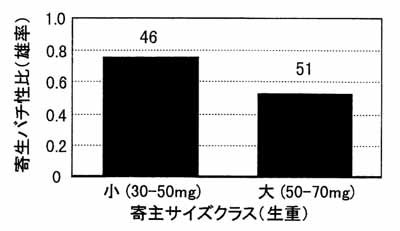
図-1キタコマユバチにおける寄主の体サイズと羽化成虫性比の関係
(数字は羽化成虫数)
それではなぜ母バチは大きな寄主に雌卵を産むのでしょうか。これには寄生バチ自体の大きさが深く関わっています。昆虫類の多くでは、体サイズが大きいほど適応度(雄の場合は交尾能力、雌では産卵能力)が高いとされています。すなわち体が大きいものほど多くの子を残せるわけです。単寄生バチの場合は、1個体の寄主がそのままハチ1個体の栄養になるわけですから、大きな寄主に寄生したハチほど大きな成虫になることができます。さらに寄生バチの場合は、体サイズと適応度との関係に雌雄間で差があるのではないか、と考えられています。これは雌のサイズと産卵数の関係が明らかな正の相関を示すのに対し、雄サイズと交尾能力との相関は雌ほど強くないということであり、体が大きくなることによって得られる利益は雄より雌の方が大きいといえます。したがって母親は大きな寄主に雌卵を産み付けるというのが、性比調節の進化に関する理論的な説明です。実際にキタコマユバチで、寄主のサイズとそれを食べて羽化したハチのサイズの関係を調べると、正の相関があることがわかりました(図-2)。また、寄主サイズが大きくなっても、それを利用したハチの雄は雌ほど大きくなっていないことがわかります。これは雄幼虫の方が寄主の摂食量が少ないために生じた差であり、大きな寄主に雄卵を産んでも寄主体のかなりの部分が食べ残されるため、「無駄な」部分が多いことを示しています。さて、それでは実際にキタコマユバチの体サイズと適応度の関係には雌雄差があるでしょうか。これを実験的に確かめるのはこれからの課題です。
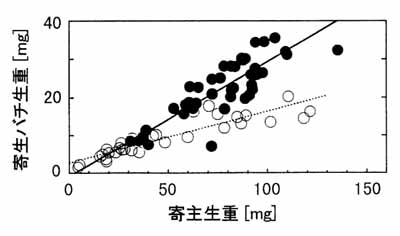
図-2キタコマユバチにおける寄主の体サイズと羽化成虫体サイズの関係
(○:♂、●:♀)
連載
似た鳥くらべ (2)
シジュウカラ・コガラ・ヒガラ・ヤマガラ
鳥獣研究室 日野輝明
ひっそりと静まり返る冬の森林にも、寒さに負けず元気良く動き回る鳥たちの一団があります。良く観察してみて下さい。いろいろな異なった種類の鳥たちが一緒になって行動していませんか。そのような群れは混群と呼ばれ、その中心メンバーとして活躍しているのが、シジュウカラ(Parus major)、コガラ(P. montanus)、ヒガラ(P. ater)、ヤマガラ(P. varius)といったカラ類と呼ばれる鳥たちです。
彼らは全長11~14cmの小さな鳥で、外見もよく似ていますが、それぞれに個性ある風貌をしており、見分けるのはけっして難しくありません(イラスト参照)。ヤマガラが日本周辺の限られた地域にしか見られないのに対して、他の3種はユーラシア大陸に広く分布します。シジュウカラとヤマガラは、平地でも公園のようにまとまった広葉樹の林があれば見ることができ、私たちにとってなじみの深い鳥たちです。一方、コガラとヒガラはある程度標高が高く、針葉樹が混じるような林でないと見られません。
いずれも枝抜けやキツツキの古巣などの樹洞を営巣場所として使い、巣箱もよく利用します。春から夏にかけては樹木の葉についているチョウやガの幼虫を主に食べ、秋や冬になると樹木の種子も食べるようになります。体は大きいものからシジュウカラ、ヤマガラ、コガラ、ヒガラの順で、けんかもこの順で強くなり、弱いものが見つけた餌を強いものが横取りするといった光景がしばしば見られます。一方、体の小さい種類は、その体を活かして枝葉に器用にぶら下がったり飛びついて、餌をすばやく探し出して食べることができます。まさに「天は二物を与えず」で、だからこそ彼らは同じ群れの中にいても一緒にうまくやっていけるのだと考えられます。

イラスト(瀬川也寸子)
おしらせ
山城試験地に二酸化炭素動態観測施設建設
森林群落の二酸化炭素吸収・放出量を観測するための施設が、京都府相楽郡山城町にある山城試験地に建設されています。施設は、森林群落と大気間での二酸化炭素の移動量などを測定する器械や、それを設置するためのタワー等から構成されています。この施設は、日本各地の森林における二酸化炭素吸収・放出量の実態を把握する目的の一環として建設されたものです。山城試験地の他に、札幌、安比(岩手県)、富士吉田(山梨県)、鹿北(熊本県)にも建設され、森林総合研究所の本支所が担当します。一連の観測から、気候変動や森林の変化によって二酸化炭素の吸収量がどのように変化するのかを予測できるようになることが期待されます。(玉井幸治)

Copyright © Forest Research and Management Organization. All rights reserved.