
ホーム > 研究紹介 > 刊行物 > 森林総合研究所関西支所年報第38号 > 年報第38号 主要な研究成果
ここから本文です。
年報第38号 主要な研究成果
1.群落レベルの物質収支特性に基づくタケ群落の管理システム
井鷺裕司・伊東宏樹・清野嘉之(造林研究室)
1. はじめに
竹林では毎年稈がタケノコという形ですばやく確実に更新する。この様な性質は一般の植林地で利用されている樹種にはないものであり、持続的な生産を極めて容易にする優れた性質として注目に値する。しかしながら、どの程度の間伐強度や間隔が特定の管理目的に対して最適であるのか、といった疑問に対してタケ群落の生態特性に基づく回答はなされていない。これまで造林研究室でタケ群落を対象に行ってきた群落レベルの物質収支に関する調査から、タケ群落の物質収支を記載するモデルを作成し、様々な管理(間伐強度、間伐間隔)に対して、タケ群落がどの様な反応を示すかシミュレーションを行った。
2. モデル
タケ群落を光合成器官である葉と貯蔵器官である根茎、そして地上部の支持器官である稈の三つのコンパートメントからなるモデルで記述した。葉で生産された光合成産物が支持器官や貯蔵器官で消費され、残りが再生産に使われる。呼吸消費速度や葉での生産効率等のパラメーターは、マダケ群落で実測された結果をもとに計算し、モデルに与えた。
3. 結果と考察
様々な間伐強度と間伐間隔の組み合わせが現存量にどの様な影響を与えるかシミュレートした結果、現実的な反応パターンが得られた(図-1 (A), (B))。間伐強度と間伐間隔には様々な組み合わせがあるが、稈の持続的な収穫量を最大にする、といった特定の管理目標を満たす間伐方法についても計算を繰り返すことで最適な間伐強度を計算できた(図-1 (C))。本モデルは成熟したタケ群落の物質収支に基づくパラメーターを用いたが、一斉開花後の回復過程についても現実のパターンにきわめてよくあてはまる計算結果が得られた。また竹林の分布拡大が都市近郊部で最近問題になっているが、刈払い等が群落に与えるダメージを評価することも可能と思われる。
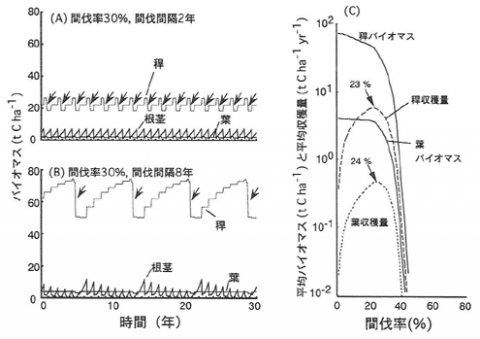
図-1 モデルの出力例
(A)間伐率30%、間伐間隔2年の時のバイオマスの経時変化。矢印は間伐を示す。(B)間伐率30%、間伐間隔8年の時のバイオマスの経時変化。矢印は間伐を示す。(C)間伐間隔2年で、間伐率を変化させたときの平均バイオマスと平均収穫量。矢印は稈と葉の収穫量を最大にする間伐率を示す。
2.萌芽が更新に及ぼす影響のモデル化
伊東宏樹(造林研究室)
1. はじめに
関西の都市近郊二次林には萌芽の性質をもつ樹種が比較的多く見られる。近年、萌芽が個体群動態にどのような意味をもつかについて考える研究が増えつつあり、撹乱との関係が指摘されている。本研究では、個体群の動態を予測する方法としてよく用いられる推移行列モデルを使って、萌芽が個体群動態に対してどのような影響を及ぼしているのかを考察した。
2. 方法
推移行列モデルとして次のような式を定義した。
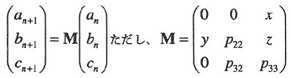
ここで、anは第n世代における種子数、bnは第n世代における非繁殖個体数、cnは第n世代における繁殖個体数である。また x, y, z はそれぞれ1繁殖個体当たりの種子生産数、種子から非繁殖個体への推移確率、1繁殖個体当たりの萌芽生産数をしめす。P22, P32, P33はそれぞれ非繁殖個体がそのままとどまる確率、非繁殖個体が繁殖個体へ推移する確率、繁殖個体がそのままとどまる確率をあらわす。このとき行列Mの固有値λは、この個体群の増殖率を示す。
既存の資料から京都東山のシイ林についてP22, P32, P33をあてはめてみたところ、
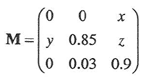
となった。ここで、 x, y, zを変化させたときにλの値がどのように変化するのかを調べた。
3. 結果及び考察
yの値を0, 0.003, 0.006と変化させたとき、x, z に対してλがどのように変化するのかを図-1に示す。これを見ると、yが小さいとき(種子があまり定着しないとき)、または x が小さいとき(種子生産量が小さいとき)に相対的に z 軸に対するλの傾きが大きくなる(萌芽が個体群動態に与える影響が相対的に大きい)ことがわかる。逆に、x,yがともに大きいときには萌芽が与える影響は相対的に小さいといえる。以上の結果は直観的にも理解しやすいものである。今後、ある林分についてのパラメータを決定することにより、その林分の更新をはかるときに、種子と萌芽のどちらによる更新を促進したほうがより効率的かなどといった問題についてもこのモデルは応用が可能と考えられる。
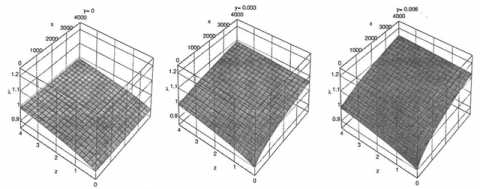
図-1 yが0, 0.003, 0.006のときのx, zに対する増殖率λの変化
3.非継続調査データに基づくスギ材積成長に対する温暖化の影響予測
杉村 乾(風致林管理研究室)
1. はじめに
スギの材積成長について、成長量と気温、降水量、純日射量、材積との間の相互関係をもとに、気候変動による生産力の変化を予測するためのシミュレーションモデルを構築し、その関係式におけるパラメーターを決定した。また、作成したモデルをもとに、温暖化の材積成長への影響予測を試みた。
2. 研究方法
日本海側の国有林収穫試験地と民有林密度管理調査地におけるスギのデータをもとに、森林動態モデル(JABOWA-II)と筑後モデルを組み合わせたモデルを作成し、パラメーターの値を決定した。密度管理調査地データ(市町村、標高、林齢、上層木樹高、材積、ha当たり本数)は継続的に測定されたものでないため、このモデルに適用するための手法を検討した。そして、GISS(ゴッダード空間科学研究所)による温暖化の予測値に気候が変動したときの生産力の変化を、北九州(福岡、佐賀、長崎)、山陰(兵庫北部~山口)、北陸(新潟~京都北部)、東北(秋田、山形)の4地域に分けて予測した。
3. 結果と考察
密度管理調査地データは、同じ林齢のプロット間でha当たり本数のばらつきが大きいので、各プロットの成長量を推定する際、上層木樹高を指標に用いる方が適当であると考えられる。そこで、各地域内でha当たり本数が平均よりも多いプロットについて、樹高と材積の関係を直線または二次曲線で回帰し、各プロットにおけるha当たり材積を推定した。また、林齢と材積の関係をロジスチック曲線または直線で回帰し、各地域の平均的な成長曲線とした。そして、以下の関係式から各プロットにおける材積成長量を推定した。すなわち、
年成長量=(プロットの材積推定値)/(プロットと同一林齢の平均材積)×(プロットと同一林齢での平均年成長量)。
ここで得られた各プロットの年成長量推定値とha当たり材積及び温量示数、成長期降水量、純放射量を用いて、以下の材積シミュレーション・モデル式におけるパラメータを両辺の対数を重回帰分析することによって決定した。
dV/dt = A*(a-V)V*(b-W)W*R*P*exp(-δ*(R/P)2)
(A、a、b 及びδは定数、V、W、R、P は材積、温量示数、水平面日射量、成長期降水量)
ここで、気候変動影響予測の際に外挿を避けるために、隣り合う2地域をまとめて計算した(表-1)。その結果、東北・北陸では成長期降水量の増加が、山陰・北九州では成長期純放射量の増加が成長量の増加につながるであろうが、温量示数の増加は材積成長にとってマイナスであることが予測された。また、乾燥の影響は山陰・北九州のみで影響が有意であると推定された。最後に、いくつかの地点を選び、現在の気候値とGISSによる温暖化予測値をモデルに代入し、現在と気候変動後の材積の変化を40年生林分について比較した結果、北陸以外の3地域では成長量の減少が予測された。これは、北陸では成長期降水量の増加によるプラスの影響が温量示数の増加によるマイナスの影響を上回ったが、他の地方では温度の上昇に比して降水量あるいは純放射量の増加幅が小さいためと解釈される。
| W | a-W | P | R | (R/P)2 | V | b-V | 定数 | ||
| 東北・北陸 | 偏回帰係数 | -0.998 | - | 0.515 | - | - | 0.657 | 0.661 | -4.73 |
| (n = 100, R = 0.843) | 偏相関係数 | -0.223* | - | 0.296** | - | - | 0.448** | 0.462** | |
| 北陸・山陰 | 偏回帰係数 | -5.19 | -6.22 | 1.66 | - | - | 0.576 | 0.771 | 35.2 |
| (n = 157, R = 0.653) | 偏相関係数 | -0.203** | 0.153* | 0.384** | - | - | 0.448** | 0.189* | |
| 山陰・北九州 | 偏回帰係数 | -2.34 | - | - | 2.51 | 912 | 0.475 | 0.510 | -1.72 |
| (n = 127, R = 0.509) | 偏相関係数 | -0.294** | - | - | 0.229* | 0.149 | 0.378** | 0.299** |
単位は℃ (W)、mm (P)、kWh/m2(R)、m3(V)。**、*は、それぞれ1%、5%有意を示す。
4.京都府上世屋・五十河地区における里山ランドスケープの特徴
深町加津枝(風致林管理研究室)
1. はじめに
我が国で薪炭林等として利用されてきた二次林やマツ林などの里山は、その周囲に立地する集落や農地とあわせて特徴的な土地利用セットを形成している。この土地利用セットである里山ランドスケープは、営林や営農にかかわる技術、政策の変化や、自然災害やマツ枯れ等の自然的変異が発生する中で、社会的・自然的要因が相互に関連し合いながら今日みられる姿に形成されてきたものである。里山を中心とした里山ランドスケープの特質を明らかにすることは、里山の分布した地域において自然環境の保全計画を策定する上での重要な基礎資料となるものと考えられる。
2. 調査方法
研究対象地は宮津市上世屋地区及び大宮町五十河地区の計約2,000haであり、典型的な過疎地域である。1970年代頃まで薪炭林利用が主体であり、スギ・ヒノキなどの造林の歴史は浅い。今日では、シデ・ナラ類等の広葉樹二次林が広大な面積で広がり、アカマツ林が主に標高400m以下の山頂や尾根筋等に小面積で点在する。また、里山ブナ林が分布する近畿地方で数少ない地域のひとつである。主な産業は稲作中心の農業で、集落周辺を中心に棚田が広がる。
用いた資料等は明治以降の5万分の1地形図、森林簿、森林調査簿、森林基本図、森林計画図であり、土地利用の分布形態の変化、1996年現在の樹種・林齢分布の空間パターンを把握した。この際、広葉樹林、アカマツ林、スギ・ヒノキ林の林齢を66年生以上、36~65年生、35年生以下の三つに区分した。そして、過去の土地被覆と1996年の樹種・林齢分布の空間パターン図を50m四方のセルに分割して比較し、今日の里山ランドスケープの特徴を考察した。
3. 結果と考察
図-1は1900年頃の土地被覆と1996年の林齢分布を含む土地被覆とを比較し、里山の変容パターンを解析した結果である。図中の数字は、各区分に含まれる解析単位であるセル数と割合を示す。各種変容パターンのうち最も数的に多いのは、1900年頃に「広葉樹」であったセルが1996年に「広葉樹・66年生以上」になったものである。ついで多いのが、1900年の「広葉樹」から、1996年の「広葉樹・36~65年生」へと変化するという、広葉樹林であった里山が、1930~1960年に伐採などのインパクトを受け、その後再び広葉樹林となったパターンである。また、1900年の「針葉樹」が1930~1960年の間に「広葉樹・36~65年生」に変化したというパターンも11%を占める。1900年に樹林地以外であったセルの中では、「荒地」から「広葉樹林・36~65年生」などのパターンが多い。
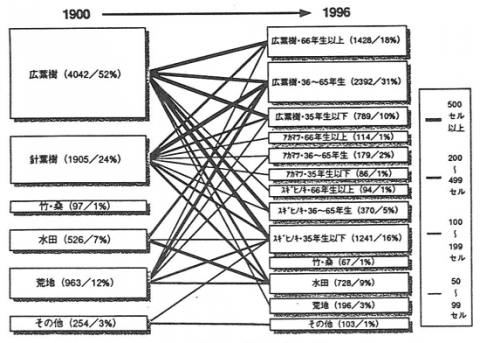
図-1 里山の変容パターン
このように、広葉樹を主体とした対象地の里山は約100年間、広葉樹林のまま遷移してきたものはわずかで、伐採の履歴があったり、水田や荒地等からの転用があった等、様々な社会的インパクトが発生していた。そして、小規模な面積単位ごとに異なった頻度や大きさで発生しインパクトにより、複雑な林相分布が形成された。以上のような過程を経て、水田、広葉樹林、アカマツ林、スギ・ヒノキ林、荒地が集落からの距離や所有形態、樹種とほぼ無関係に混在するのが今日の里山ランドスケープの特徴といえよう。
5.国土数値情報を利用した湖西地域の被視ポテンシャル解析
奥 敬一(風致林管理研究室)
1. はじめに
被視ポテンシャルを指標とした景観解析は、地域の景観にとって重要な構成要素となる地点を明らかにすることができ、とくに数km~数十kmスケールの景観計画を策定する上で基礎となる解析である。そして、これに土地の被覆状況を併せることで、地域の景観特性を把握することが可能となる。広域的な観点からこのような景観特性を考える際に土地被覆状況を表現する情報としては、国土数値情報が利用できる。土地利用や植生については約1km四方の三次メッシュ(以下メッシュ)が、数値地形データとしては三次メッシュを16分割した250mグリッド(以下グリッド)の標高値が、日本全国について利用可能である。本論では、これら国土数値情報を利用して被視ポテンシャル解析を行うことにより、流域レベルにおける農林地の景観機能解析への応用を試みた。
2. 方法
対象地域は琵琶湖集水域の西側に当たる滋賀県湖西地域(1/25,000地形図、饗庭野、今津、北小松、勝野、比良山、沖島の滋賀県内の範囲)であり、メッシュ数595、グリッド数9,520(うち陸域5,385)である。各メッシュは国土数値情報の土地利用面積割合をもとに、図-1のフローに従って7種の景観タイプに類型区分した。景観タイプは視対象としての土地被覆を表現するとともに、視点場としての容量(たとえば人の多さなど)も表すものとし、表-1のように視点ウェイト値を与えた。ただし、このウェイト値は日常的な生活・行動範囲を念頭に置いて数値を与えたものであり、非日常的な利用に伴う視点場については考慮しないものとした。また、土地利用面積割合の土地利用区分では樹林地はすべて森林として一括されているため、景観タイプだけからでは農林地の植生の状況が把握できない。そこで、三次メッシュ現存植生データから植生自然度(以下自然度)の項目を利用し、植生の状況を補足した。
250mグリッドのある一点(座標(x,y))の被視ポテンシャル値Vxyは次のような式で表される。
![]()
Wi: 景観タイプiの視点ウェイト
Ji(u,v): 景観タイプ がiの座標(u,v)と(x,y)の間の可視判定関数、可視ならJ=1、不可視ならJ=0
D: 座標(u,v)と(x,y)の間の視距離判定関数、7km以内ならD=1、7kmを超えるとD=0
視距離判定関数において7kmを境界としたのは、既存の研究における樹冠テクスチャの弁別限界としてあげられているためである。
以上から、山間部に該当する景観タイプ、及び自然度ごとにV値を集計し、対象地域における農林地の景観特性を考察した。
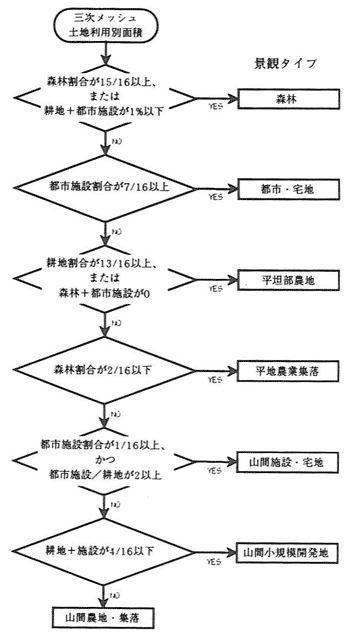
図-1 景観タイプ区分フロー
| 景観タイプ | 視点ウェイト値 |
| 都市・宅地 | 1.0 |
| 平坦部農地 | 0.1 |
| 平地農業集落 | 0.6 |
| 山間施設・宅地 | 0.4 |
| 山間農地・集落 | 0.3 |
| 山間小規模開発地 | 0.2 |
| 森林 | 0 |
3. 結果と考察
表-2には山間部について景観タイプ、自然度ごとのV値の集計結果、グリッド数を示した。自然度は森林植生及び、田畑について記した。
景観タイプごとの特徴をみると、V値の合計は森林景観タイプがもっとも大きく、山間農地・集落景観タイプがこれに次ぐ。湖西地域は湖岸の低地から山地上部までの標高差が大きく、平野から台地部にかけての狭い地域に市街地、集落、農地が集まり、その上部には森林が広がる。そのため地域全体としては森林景観が卓越し、斜面下部に農地や集落などのセットである農村景観が広く混交するような景観が見られやすいという特徴を表している。1グリッド当たりのV値では山間農地・集落景観タイプが高い値を示すが、この景観タイプに設定された視点ウェイト値が高いことに加え、市街地に近接していることが影響していると考えられる。
自然度についてみると、二次林と田畑のV値合計が大きな値を示す。二次林、田畑のグリッド数が非常に多いこともひとつの理由であるが、植林地や自然林に比べても1グリッド当たりのV値は大きい。とくに樹林地に関して言えば、集落や市街地近郊の森林の多くが二次林であって、その視覚的な重要性が高いことを示唆している。湖西地域の景観を構成する主要な要素と考えられる山間農地・集落景観タイプの中でも、自然度では二次林として区分されるグリッドが4割弱存在し、1グリッド当たりのV値も大きい。集落、農地、二次林のセットであるこうした景観は、農業技術や農林業構造の変化に伴い、今後の変質が予想されているが、現代においても視覚的に重要な位置を占めているといえる。自然林についてはグリッド数自体も少ないが、視点場としての容量が大きい地域から遠く、視覚的に隔離された立地にあると考えられる。
今回得られた結果から、国土数値情報を使用した被視ポテンシャルの解析が、流域スケールの景観特性把握に有効であることが示された。今後、広域の景観計画策定に際しては、地域における主要な景観タイプが将来どのような推移をたどり、またどのように誘導すべきかを考えていく必要があろう。
| 植生自然度 | |||||
| 田畑 | 植林地 | 二次林 | 自然林 | 農林地計 | |
| 被視ポテンシャル値(V) | |||||
| 山間施設・宅地 | 0 | 609.9 | 5387.5 | 0 | 5997.4 |
| 山間農地・集落 | 22017.0 | 381.8 | 12834.5 | 2622.8 | 37856.1 |
| 山間小規摸開発地 | 314.9 | 3914.9 | 19467.4 | 0 | 23697.2 |
| 森林 | 0 | 540.4 | 57227.5 | 1289.8 | 63921.7 |
| 山間部計 | 22331.9 | 10311.0 | 94916.9 | 3912.6 | 131472.4 |
| グリッド数(G) | |||||
| 山間施設・宅地 | 0 | 16 | 130 | 0 | 146 |
| 山間農地・集落 | 423 | 15 | 262 | 16 | 716 |
| 山間小規摸開発地 | 32 | 205 | 536 | 0 | 773 |
| 森林 | 0 | 284 | 2052 | 144 | 2480 |
| 山間部計 | 455 | 520 | 2980 | 160 | 4115 |
| V/G | |||||
| 山間施設・宅地 | 38.12 | 41.44 | 41.08 | ||
| 山間農地・集落 | 52.05 | 25.45 | 48.99 | 163.93 | 52.87 |
| 山間小規摸開発地 | 9.84 | 19.10 | 36.32 | 30.66 | |
| 森林 | 19.03 | 27.89 | 8.96 | 25.77 | |
| 山間部計 | 49.08 | 19.83 | 31.85 | 24.45 | 31.95 |
二次草原は現存植生データの自然度4および5を合わせたもの
二次林には「自然に近い二次林(現存植生アータの自然度8に該当)」も含む
6.モウソウチクとマダケの植物珪酸体含有率
鳥居厚志(土壌研究室)
1. はじめに
イネ科など特定の植物分類群に多く蓄積されている植物珪酸体(plant opal;以後POと略す)に関して、これまで多くの報告がなされているが、タケ類については詳細な基礎データの蓄積が少ない。竹林の分布変化に関する研究の一環として、タケ類の植物珪酸体含有率を調べた。
2. 研究方法
一般に植物種を問わず、POはそのほとんどが葉の表皮細胞で生成される。しかしタケ類は、① 一般的なイネ科等の草本に比べて、葉以外の器官のバイオマスの比率が高く、② 稈や鞘も緑色を呈し光合成を行っている。そこで葉以外の器官でも無視できない量のPOが生産されている可能性があると考え、器官別にPO含有率を調べた。
供試したのは、日本で分布面積の広いモウソウチク(Phyllostachys pubescens)とマダケ(Phyllostachys bambusoides)である。採取場所は森林総研関西支所構内(京都市伏見区)、島津実験林(京都市伏見区)、木津試験地(京都府木津町)で、葉、葉鞘、稈、稈鞘、枝、地下茎、根に分けて採取した。採取時期は、葉と葉鞘は落葉期、稈鞘は稈が伸長して脱落する時期である。それ以外の器官は春に、発生後5年以上の稈とその地下部から採取した。試料は、乾式灰化(磁性坩堝で燃焼)-塩酸洗浄-重液分離(臭化亜鉛: 比重≒2.28)を行い、POを回収して含有率を調べた。
3. 結果と考察
表-1にPO含有率の分析結果をまとめた。器官別のPO比率をみると、マダケ、モウソウチクとも葉が70~80%を占めるが、それ以外の器官でも無視できない量のPOが生産されている。葉以外では葉鞘に多い。地下部からも多量の珪酸物質が検出されたが、珪酸が最終的に地下部の細胞に沈着するかどうかは不明なので、器官別の比率の計算からは除外した。なお表をみる限りでは、マダケのPO含有率はモウソウチクのそれの2倍程度(葉)であるが、両種の試料採取場所の立地条件が異なるので(マダケ; 洪涵地、モウソウチク; 丘陵地)、一概に種の特性による差であるかどうかはわからない。
回収したPOを顕微鏡で観察した結果は、形態的にササ類やネザサ類との共通性が目立った。すなわち大型粒子の中ではFan I が多く、Fan/Elongate 比が大きい。また小型粒子の中では Bambusoid が多いが、ササ類に多い Bambusoid Iよりもネザサ類に多い Bambusoid II の方が主であった。Chloridoid もある程度含まれていた。ただし葉以外の器官から回収されたPOは小型の不定形のものが多く、通常の形態分類体系では分類不可能であった。これは細胞への珪酸の沈着が十分ではなく、POとしては未熟な粒子であるためと思われる。この点を考えると、葉以外の器官で生産されたPOは、量的にはある程度の意味はあるが、古植生の復元目的などのためには重要性は小さいといえる。
| 種 | 部位 | 絶乾試料重(g) | 回収したPO重(g) | PO含有率(%) | 器官別のPO比率(%) |
| マダケ | 葉 | 13.54 | 1.664 | 12.29 | 78.1 |
| 葉鞘 | 7.02 | 0.651 | 9.27 | 16.7 | |
| 桿鞘 | 11.23 | 0.157 | 1.40 | 1.1 | |
| 桿 | 20.47 | 0.033 | 0.16 | 1.3 | |
| 枝 | 18.25 | 0.321 | 1.76 | 2.8 | |
| モウソウチク | 葉 | 3.62 | 0.227 | 6.27 | 72.6 |
| 葉鞘 | 4.39 | 0.172 | 3.92 | 19.8 | |
| 桿鞘 | 11.89 | 0.171 | 1.44 | 1.4 | |
| 桿 | 33.49 | 0.046 | 0.14 | 1.4 | |
| 枝 | 34.74 | 0.342 | 0.98 | 3.6 | |
| (根) | 9.90 | 0.088 | 0.89 | — | |
| (地下茎) | 9.66 | 0.078 | 0.81 | — |
根と地下茎は、最終的に珪酸が沈着するかどうか不明なので,括弧書きで示す.
7.土壌発達の初期過程―土壌微生物バイオマスCの変化―
古澤仁美(土壌研究室)
1. はじめに
治山施工後の植生回復に伴う土壌の変化を明らかにすることは、目的とする森林が回復するかどうかを予測し、目的を達成できる施工方法を確立するために必要である。そこで、施工直後からの植物の成長と土壌の化学性、物理性の変化を調査することにした。今回は、そのうち微生物バイオマスCの推移について初期1年間の結果を報告する。
2. 方法
関西支所構内実験林の斜面(傾斜42~45°、北西向き)に、S1:アカメガシワ区、S2:ヤシャブシ区、S3:アカマツ区、S4:草本区、S5:対照区(裸地)の5つの試験区を設けた。各試験区にマサ土を厚さ6cmに客土し、各々の種子の期待本数密度を木本: 200本/m2、草本: 4,000本/m2に設定して1996年4月に播種した。施工後、散水は表面が乾かない程度に適宜行った。各区の成立本数、成長量の測定とともに、表層土壌(深さ0~5cm)の微生物バイオマスCをクロロホルム薫蒸抽出法を用いて2ヶ月毎に測定した。
3. 結果と考察
成立本数は施工から7ヶ月後の11月でアカメガシワ区、ヤシャブシ区、アカマツ区、草本区でそれぞれ1.6本/m2、28本/m2、352本/m2、648本/m2であった。
11月までの成長量(上位10本の平均)はアカメガシワ区、ヤシャブシ区、アカマツ区、草本区でそれぞれ 33.7cm、2.0cm、5.6cm、50.8cmであった。アカメガシワは成立本数が少ないが、成立した個体は他の木本種に比べて速い成長を示した。
微生物バイオマスCは、施工後1年間では樹種の違いによる差は認められなかった。いずれの区でも施工1ヶ月後の5月では0~50μg/g soil 以下であった。施工から7ヶ月後の11月には80~150μg/g soilに増加し、その後は全ての区で100μg/g soil 以上で一定している(図-1)。同時に測定した関西支所構内の常緑広葉樹林では1年間の平均が2,376μg/g soilであり、試験地をそれと比較すると1オーダー以上小さかった。微生物バイオマスCは土壌の有機物含量に比例して増加するといわれている。構内常緑広葉樹林の炭素、窒素含有率が13.93%、0.86%であるのに比べ、施工時における試験地ではそれぞれ0.46%、0.005%であり、非常に乏しかった。現在試験地では、バイオマスCは有機物含量と平衡状態になりつつあると考えられるが、今後土壌有機物の蓄積と微生物バイオマスCの推移を長期的に見ていく必要がある。
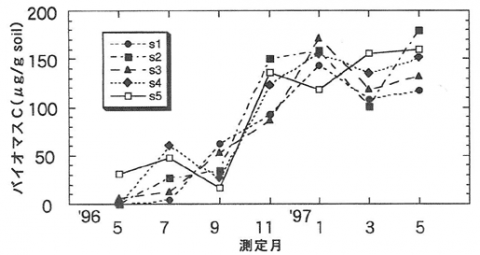
図-1 緑化試験地でのバイオマスCの推移
8.大台ヶ原の森林及び土壌の状況
金子真司・古澤仁美・鳥居厚志・荒木 誠(土壌研究室)
西本哲昭(関西支所)
伊東宏樹(造林研究室)
1. はじめに
大台ヶ原は近畿地方の中で亜高山帯針葉樹林が分布する数少ない地域であるが、伊勢湾台風(1959年)に端を発するトウヒ林の衰退に加え、シカの食害による衰退が進みつつあり、問題となっている。現時点では酸性降下物の直接の影響はないとみられるが、亜高山帯針葉樹林の分布の限界に位置していることに加え、年降水量がおよそ5,000mmと非常に厳しい気象条件下にあるために、環境の変化に対して非常に弱い立場にあると推定される。この大台ヶ原の樹木及び土壌の状態について調査を行った。
2. 方法
大台ヶ原の牛石が原近くの針葉樹林(N34° 10’ 12”、E136° 6’ 13”)において、平成8年9月11~12日に酸性雨調査マニュアル1)にしたがって樹木の成育状況及び土壌の状態について調査を行った。
3. 結果及び考察
平均樹高は14.6m、平均胸高直径は35.6cmであり、胸高直径に比べて樹高が低く、調査木の30%が枯死木であった。林床に生育するイトザサ(ミヤコザサ)はシカの食害によって丈の高さが15cm程度にそろっていた。土壌型は適潤性褐色森林土(BD)であったが、A~B層の腐植含量は高く、暗色系褐色森林土に近い断面形態をしていた。A層とB層は礫が少なかったが両層をあわせても26cmと薄く、C層は巨礫を多量に含んでいた。表層土壌16地点の平均pHは4.39±0.18で、EC (電気伝導度)は2.88±0.9mS/m であった。図-1に示すように、土壌水中のイオン濃度はH+以外のすべてのイオンがスギ林土壌(1996年9月18日調査)に比べて濃度が低かった。特にCa2+やMg2+が少ないことは、硝化によって生成したNO3-を中和する塩基類が不足していることを示しており、そのためにpHの低下がおきているものと推察された。
以上のように大台ヶ原の森林は樹高が低く、枯死木が多いという特徴がみられたが、その原因には寒冷な気候に加えて、有効土層が薄いことや土壌の酸性が強く塩基が乏しいことなどが関係していると考えられた。すわなち、大台ヶ原の土壌は現在でも塩基が乏しく低pHであるが、酸性雨の影響をうけると塩基の溶脱が進み土壌はさらに酸性化するために、樹木の生育に大きな影響を与えると予測された。そこで、大台ヶ原の森林及び土壌については今後もモニタリングを行っていく必要があると考えられた。
図-1 土壌中の水溶性イオン濃度(meq/L)の比較
1)林野庁 (1990) 酸性雨等森林被害モニタリング事業実施マニュアル.
9.ヒノキ人工林における土壌炭素貯留量
荒木 誠・金子真司・鳥居厚志・古澤仁美(土壌研究室)
西本哲昭(関西支所)
1. はじめに
全世界の森林土壌中には、大気中の全炭素量に匹敵する6~7千億トンもの炭素が固定されている。これらの炭素は、森林が成立し、成熟していく過程で徐々に固定されてきたものと考えられる。そこで、森林の炭素貯留量を評価していくためには、土壌中の炭素貯留の実態を把握するとともに、土壌炭素貯留量の推定を行うことが必要である。ここでは、人工林施業における土壌炭素貯留量を解明する一環として、ヒノキ人工林の土壌調査・分析の結果を用いて土壌炭素貯留量と土壌特性の関係を解析し、土壌炭素の貯留量を解明するとともに推定方法を検討した。
2. 研究方法
関西地域のヒノキ人工林において土壌調査・分析を行うとともに、既往の中国地方、中部地方、及び関東地方のヒノキ林の土壌調査結果をもとに、土壌の諸特性と土壌炭素含有率との関係を解析する。
3. 結果と考察
99箇所のヒノキ人工林の調査結果から、土壌調査を行った深さ(通常1m深以下)までの土壌炭素貯留量を求めた。その結果、林分における地形、地質、土壌などの立地環境条件や林木の成育状況などによって様々な値を示したが、150~200t・ha-1の林分が最も多く、大部分のものが 100~250t・ha-1の範囲にあった(図-1)。
土壌の諸特性と土壌炭素含有量との関係を解析したところ、炭素含有率と細土の容積密度(単位体積当たりの細土の質量)との間に(1)式に示す関係が見い出された。
| log BD =-0.281・log C - 0.0277 | ……… | (1) | ||
| (BD: 土壌の容積密度、C: 土壌の炭素含有率) | ||||
一方、層位ごとの単位面積・単位層厚当たりの土壌炭素貯留量は、容積密度と土壌炭素含有率の積で求められるので、(2)式が得られる。
| Scui= Ci・10(-0.281・log Ci - 0.0277) | ……… | (2) | ||
| (Scui: 層位iの単位面積・単位層厚当たりの土壌炭素貯留量、Ci: 層位iの炭素含有率) | ||||
調査・分析によって得られた実測値と(2)式で得られる推定値を図-2に示した。この関係を用いれば、土壌炭素含有率が既知の場合、あるいは何らかの手段で推定できれば、任意の深さまでの土壌炭素貯留量を推定することが可能である。
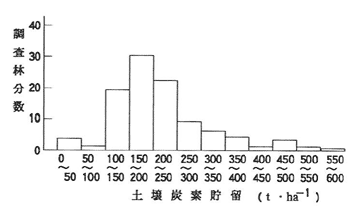
図-1 土壌炭素貯留量の頻度分布
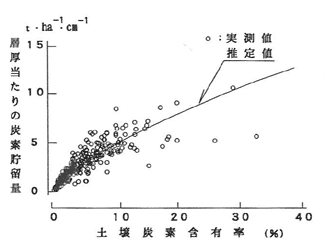
図-2 土壌炭素含有率と炭素貯留量の関係
10.京都府南部の落葉広葉樹二次林及びアカマツ林における林床可燃物量の季節変化
後藤義明(防災研究室)
1. はじめに
落雷により直接樹冠や樹幹部から発火して火災となることの多い北欧や北米大陸の諸国とは異なり、我が国の林野火災のほとんどは人為的な原因によって発生している。しかも、その多くはタバコの投げ捨て、たき火の不始末など人間の不注意に起因している。したがって我が国では、林野火災は地表部分、すなわち林床可燃物から発生することが圧倒的に多く、地表火から樹幹火や樹冠火、地中火に移行するのが普通である。林床可燃物の中でも、林床に堆積した落葉・落枝類が最も危険で、林野火災の出火源あるいは媒介物として火災延焼に重要な役割を果たしている。そこで京都府南部のアカマツ林及び落葉広葉樹二次林において、林床可燃物として重要な落葉・落枝量と、林分構造や微地形、あるいは季節の違いとの対応関係を調べた。
2. 調査地と調査方法
調査は京都府相楽郡山城町にある山城森林水文試験地北谷試験流域で行った。調査区内に3本の調査ラインを設け、各ラインにおいて5mおきに調査プロットを設置して、5m×5m枠内の胸高直径3cm以上の立木の胸高直径を測定した。また、この枠のほぼ中央に50cm×50cmの小枠を置いて枠内の落葉・落枝を採集し、その乾燥重量を測定した。さらに小枠の周辺で落葉・落枝の堆積深を測定した。乾燥重量及び堆積深の測定はL層とF+H層に分けて行った。京都市山科区御陵大谷町の林野火災跡地に設けた試験地に隣接したアカマツ林において、3箇所の調査プロットを設け、同様の調査を行った。調査は2月、5月、8月の3回行った。
3. 結果と考察
落葉広葉樹の優占する山城水文試験地での調査プロットの総数は42地点であった。全調査プロットでの落葉・落枝量の平均値は、L層がヘクタール当たり 5.9t→5.0t→3.9tと次第に減少したのに対し、F+H層はヘクタール当たり 5.2t→5.8t→6.5tと徐々に増加した。全体では11.1t→10.8t→10.4tとわずかに減少する傾向を示した。L層の深さとL層の重量との関係には有意な相関(p<0.01)が得られたが、F+H層の深さとF+H層の重量との間には有意な相関関係は得られなかった。
アカマツの優占する山科区御陵大谷町の試験地での調査結果では、L層がヘクタール当たり8.6t→6.9t→5.1tと減少したのに対し、F+H層は7.6t→9.2t→11.0tと増加した。結果として全体では、16.2t→16.1t→16.1tと全期間を通してほぼ一定の値となった。山科試験地(アカマツ林)での値は山城試験地(落葉広葉樹二次林)での値の約1.5倍あった。
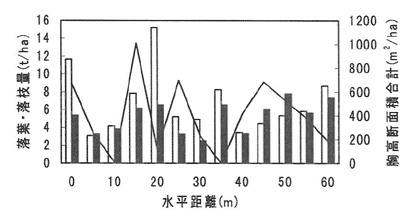
図-1には山城試験地に設けた調査ラインでの、林木の胸高断面積合計と落葉・落枝量の関係の一例を示した。胸高断面積合計の大きいところでは落葉・落枝量も大きくなる傾向があるが、その関係は明瞭ではなかった。地形と落葉・落枝量との関係に関しても明瞭な関係は見いだせなかった。
|
図-1 胸高断面積合計と落葉・落枝量との関係
|
11.落葉広葉樹二次林における「蒸発効率」の特性
玉井幸治(防災研究室)
1. はじめに
多様な植物群落の蒸発散特性を比較する尺度のひとつとして、近藤ら(1992)が提案した蒸発効率(β)がある。渡辺(1994)は、「土壌水分や植物生理だけの影響を見るためにはの値を調べる方がよい。」とし、多様な植物群落におけるの値を報告している(表-1)。それによると、水分補給が豊富な植物群落ほどの値が大きい傾向が認められる。しかし森林におけるの知見はきわめて限られたものであるため、事例を増やす必要がある。そこで本報告では、京都府南部の寡雨乾燥地帯に位置する落葉広葉樹林におけるの季節変化を推定し、既往報告との比較を行った。
2. 理論
近藤ら(1992)によると、植物群落からの顕熱フラックス(H)、潜熱フラックス(lE)は、(1)、(2)式によって表される。
H = CP ρ CH U (TH - Ta) |
(1) |
lE = l ρ β CH U (qSAT (TE) - qa) |
(2) |
CP: 空気の低圧比熱, ρ: 空気の密度, CH: 顕熱のバルク輸送係数, U: 風速, β: 蒸発効率, TH: 顕熱, TE: 水蒸気に対する有効地表面温度, Ta: 大気温度, qa: 比湿, qSAT: 飽和比湿, l: 水の気化熱
| Rn - G = H + lE | (3) |
Rn: 純放射量、G: 地中熱流量
で表される熱収支式に(1)、(2)式を代入し、TH=TEと近似することによって
Rn - G = CP ρ CH U (TE - Ta) + l ρ β CH U (qSAT (TE) - qa) |
(4) |
が導出される。(4)式は、TEと1Eが未知数であってもRnなどの観測値を与えて逐次近似法を用いることにより、任意のに対しlEを求めることができる(渡辺、1994)。
3. 観測林分と方法
その時々の観測値に応じてとlEの関係は一義的である。そのためlEの値が既知であれば、逆にの値を求めることができる。本研究ではThornthwaite-Holtzmanモデルまたはボ-エン比法により算出し、短期水収支法によって検証を受けたlEの値(Tamai &amp; Hattori、1994)を用いて、2つの未知数であるとTEの値を解とした。
対象林分は京都府相楽郡山城町北谷国有林にある山城森林水文試験地北谷流域(北緯34°47’、東経135°51’)である。主林木はコナラ、ネジキなどの落葉広葉樹である。胸高断面積合計は常緑樹6.29m2/ha、落葉樹13.31m2/haと比較的小さな値である。Rnなどの森林微気象観測は、流域内の尾根地形上に設営された気象観測鉄塔に取り付けられた測器によって行った。気象観測鉄塔周辺の平均樹高は約6mであった。測器とその測定高は、純放射計(英弘精機製CN-11): 10m、通風筒付き乾湿球温度計(英弘精機製MH-020): 8、10m、3杯式風速計(中浅測器製A701): 8、10mとした。地中熱流板(英弘精機製CN-81)を0.05mの深さに埋設した。(4)式中のTHを0.077と推定し、(4)式の計算に用いた。なお遮断蒸発は土壌水分や植物生理の影響をほとんど受けないため、降雨時の観測デ-タは解析から除外した。
4. βとTE - Taの特性
ばらつきが大きいものの、の値には季節変化が認められた(図-1)。3、4月には、0.03前後の値を示す日が最も多かった。5、6月中は葉の展開につれての値は次第に増加していき、7月前半には0.18前後の値を示した。7月後半から8月にかけては一転して減少傾向になった。この減少傾向は一時的なものか否かについて、さらに事例を増やして検討を加える必要があろう。観測期間を通じて降雨のあった日や 曇天日にはの値が大きくなる場合もあった。季節別の平均値は、着葉期(6~8月)は0.15、落葉期(3~5月)は0.06であった。渡辺(1994)が森林における値として報告している0.26(夏)、0.1(冬)に比べると本研究による値は小さい。本研究が対象とした森林は寡雨乾燥地帯に位置し、胸高断面積も少ない。そのため森林土壌が一般の森林に比べて乾燥傾向にあり、が小さくなった可能性が考えられる。この点については森林土壌水分環境の観測を行うなどして、検証を行う必要がある。
| 植生 | β |
| 水田 | 0.5~0.8 |
| リンゴ園 | 0.3~0.4 |
| 牧草地 | 0.2~0.4 |
| 森林(夏) | 0.26 |
| (冬) | 0.1 |
算出された温度差(TE-Ta)の季節変化を図-2に示す。最大値は約1.3℃であった。0.3~0.9℃の範囲を示した日が全体の68%を占めた。このことから北谷流域の温度差は一般におおよそ1℃以下といえる。5~8月にかけて、温度差が減少していく傾向が認められる。その原因には5月以降は次第に蒸散が盛んになり、葉から蒸発熱が奪われる効果が卓越するためと考えられる。Cleugh & Dunin(1995)は、コムギ畑のキャノピ-温度と気温の差について日変化を示している。それによると、キャノピ-温度の測定方法によって違いはあるものの、温度差の日変化は1.5~5.0℃程度をピ-クとする放物線的な変化であった。本研究での温度差は日平均値に相当するものであり、Cleugh & Dunin(1995)の値と比較しても妥当な値であると思われる。
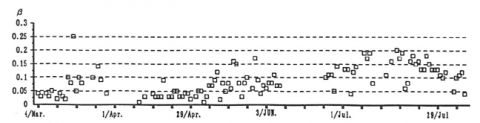
図-1 蒸発効率(β)の季節変化(92年3~8月)

図-2 有効地表温度(TE)と気温(Ta)の差の季節変化(92年3~8月)
引用文献
- Cleugh, H. A. and Dunin, F. X. (1995) Modelling sensible heat fluxes from a wheat canopy: an evaluation of the resistance energy balance model, J. Hydrology, 164, 153–170.
- 近藤純正、渡辺 力、中園 信 (1992) 日本各地の森林蒸発散の熱収支的評価, 天気,39, 685–695.
- Tamai, K. and Hattori, S.(1994) Characteristics of canopy conductance of a deciduous broad-leaved forest, In: Proceedings of the International Symposium on Forest Hydrology, 147–154.
- 渡辺 力(1994) 植物と大気, (近藤純正編著, 水環境の気象学, 朝倉出版), 208-239.
12.四つ子川における浮遊物質組成の季節変動
深山貴文(防災研究室)
金子真司(土壌研究室)
1. はじめに
河川の影響する沿岸域の生産力(プランクトン濃度)は外洋に比べ著しく高いことが知られている。また、これまでに森林が沿岸域生態系へ与える影響については特に関心が持たれており、漁場形成のために植樹運動も行われてきた。森林からは渓流を通じて浮遊有機物や微細土砂が流出しており、これらは渓流内や下流域の生態系に対して影響を与えていると考えられている。これまで浮遊物の流出形態や流出量については、北海道南部における夏期の観測例があるが、年間を通じて観測された例は少ない。そこで本研究では浮遊物質組成と浮遊物質濃度の季節変動についての観測を行った。
2. 調査地と方法
調査地は琵琶湖の西方約2kmに位置する滋賀県志賀町の四ツ子川支流と本流である。以下それぞれ支流、本流と表記する。支流の流域面積は約7.4ha、本流の流域面積は約100haである。調査地の地質は花崗岩で、主な植生は若齢スギ・ヒノキ混交林だが渓流沿いにはハンノキ、ヒメヤシャブシ等の落葉広葉樹からなる渓畔林が発達しており、渓床に堆積する落葉も主にこれらが占めている。調査期間は1996年1月から翌年1月までの1年間とした。渓流水はほぼ15日間隔で平水時に支流と本流から採水した。浮遊物の組成は、ふるいを用いて粒径別に粗粒浮遊物(粒径106μm以上2mm未満の浮遊物)と細粒浮遊物(粒径1μm以上106μm未満の浮遊物)に分類し、さらにそれらをマッフル炉によって500℃で30分間加熱し、灼熱減量から浮遊有機物量を測定した。粗粒浮遊物の測定には30L、細粒浮遊物の測定には5Lの渓流水を使用した。また、浮遊物はガラス繊維濾紙を用いた吸引濾過によって集めた。
3. 結果と考察
調査の結果、支流の浮遊物濃度は平均2.46mg/L、本流の浮遊物濃度は平均5.03mg/Lと支流より高かったが、浮遊物中に有機物が占める割合は支流で平均34%、本流で平均11%と支流より低かった。また、有機物の割合は初夏に連続してやや高くなることがあった。浮遊有機物濃度の季節変動については、4月から7月にかけて高まる現象が観測された(図-1)。さらに浮遊有機物について、FOSS(細粒浮遊有機物)とCOSS(粗粒浮遊有機物)の比であるFOSS/COSSの季節変動を調べると、夏期に粒径の細かな浮遊有機物の比率が高くなる現象が認められた(図-2)。これまでに渓流における有機物の分解については冬から春は水生昆虫、春から夏はバクテリアの活動が顕著になることが知られている。浮遊有機物成分の変動は、渓畔林から供給された落葉などが気温、水温の上昇に伴い、水生昆虫やバクテリアによって破砕、分解され次第に細分化されて流出したことによる影響の可能性が考えられた。
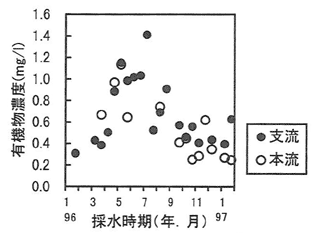
図-1 浮遊有機物濃度の季節変動
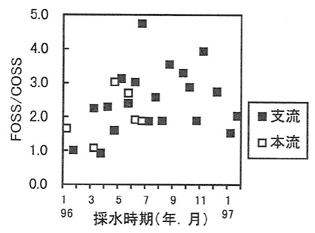
図-1 FOSS/COSSの季節変動
13.木質内装材の生産と流通
―内装用集成材のケース―
野田英志(経営研究室)
1. はじめに
健康で快適な居住環境に対する生活者(住まい手)の関心が高まる中で、居住空間を構成する内装箇所への木質素材の利用が次第に広がりを見せている。その利用は、建築物が木造か非木造か、住宅か非住宅かを問わないため、今後、大きな木材需要の場を生み出す可能性を持つ。このため、木材の供給システムを考える際には、従来の「柱材などの住宅用構造材」に重点を置くだけでは不十分であり、今後は新たに内装材分野をも考慮に入れて行くことが重要となろう。
木質内装材は一般に、他の建築用製材品に比べて、より高次加工され付加価値を高めた商品が多い。その使用箇所・形状は多岐にわたり、その生産・流通も独自のシステムをとる場合が多い。そこで本稿では木質内装材のうち、近年、増加傾向にある内装用途の集成材(造作用集成材及び化粧ばり構造用集成材)の生産・流通の仕組みを探った。
2. 内装用集成材の生産・流通
内装用集成材の平成8年の生産量は48万m3で、同年の集成材・総生産量72万m3の67%を占める(日本集成材工業協同組合調べ)。造作用集成材の生産は、高度経済成長期の昭和40年代とバブル期以降に大幅に増加している。集成材工場数は、平成7年末現在で全国に379工場あり(木材流通構造調査報告書)、北海道の48工場をトップに、奈良県41工場、秋田県26工場の順で、地域的に偏在している。平成7年の集成材製品の生産量上位5県は、奈良県が11.2万m3、秋田県7.1万m3、北海道6.6万m3、岐阜県5.0万m3、岡山県4.9万m3であり、この5道県で全国生産の51%を占めている。
図-1は、内装用集成材の流通機構を示したものである。これは、(財)日本木材総合情報センターが行った平成7年度・木質内装材流通実態アンケート調査結果表を利用し、一部推計を加えて作成した。図中の数値はJAS認定を受けた内装用の集成材製造工場112工場分の集計値である。以下、製品販売面に絞ってその特徴を摘記したい。販売ルートは、流通業者経由で販売されるものが57%と過半を占める。そのうち木材建材卸売業者(問屋)への販売が33%、商社(建材専門商社等)が14%である。製品市場・木材センターへの流通ウェイトが高い国産材製材品とは大きく異なる。建築業者への直販は34%を占めており、そのうちハウスメーカーが16%と高いウェイトを持つ。これは大手・中堅の集成材メーカーを中心に、ハウスメーカーへの直販指向が高まっているためで、その過程で、既存の流通段階が担っていた商品の品揃え機能が、次第に集成材メーカーにも求められるようになっている。このため内装材だけでなく、さらに構造材をも加えた、住宅部材総合供給企業ないし企業グループへの方向が、直需への対応力を高める上で今後重要となっている。なお、内装用集成材メーカーの中には、ホテルや各種店舗などの特殊内装部材の生産を専門とするメーカーも見られ(一般に小規模)、特注による単品生産・流通(建築現場直送)の形態をとる。このように内装用集成材の生産・流通は、規格製品(和室内装化粧材や階段セットなど)の量産・大ロット流通から、特殊材の単品生産・流通まで、多様な生産・流通形態が重層して成り立っていることがわかる。今後さらに、輸入集成材との競合関係、販売ルート間の価格形成機構の解明が必要である。
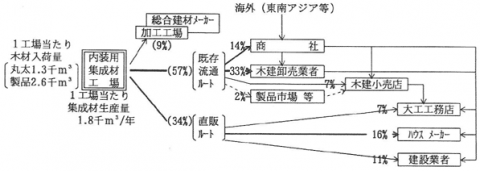
図-1 内装用集成材の流通機構(平成6年次・推定)
14.マツ材線虫病の進展と水プロトンNMR緩和時間との関係
池田武文(樹病研究室)
1. はじめに
樹木におこる病気の発病機構や抵抗性機構の解明に際しては、病原微生物の侵入に対する樹木の反応を生理学的にとらえることが重要である。そこで、植物の生命活動にとって水は不可欠の物質なので、この水の様々な挙動をとらえることは、樹木の生理状態を探る有効な手段となりうる。そのひとつとして、水プロトンNMR(核磁気共鳴)緩和時間を取り上げた。生体内の水は生体高分子や細胞の膜系との強い相互作用によって、普通の水とは異なり運動が束縛されて構造化した状態にあるものと考えらている。水プロトンNMR緩和時間(縦緩和時間(T1)及び横緩和時間(T2))は、このような生体内における水分子の運動状態を強く反映し、組織や細胞の生理的変化を調べる上で重要なパラメータとなっている。パルスNMR分光法による水プロトンの緩和時間の測定は、温度や虫害等のストレスに対する初発応答を知るための有効なパラメータであることが知られているが、樹木に対してこの方法を適用した報告は少ない。
マツ材線虫病に罹病したマツが示す代表的な病徴のひとつとしてマツの水分状態の急激な低下があげられる。その低下がマツのどのような生理的変化を表わしているのかを知る手がかりとして、マツノザイセンチュウを接種後の水プロトンNMR緩和時間を測定した。
2. 材料と方法
4年生クロマツ鉢植え苗の1年生枝に1万頭のマツノザイセンチュウを接種し、その後の枝と針葉の水プロトンNMR緩和時間ならびに含水量を測定した。
3. 結果と考察
接種後、針葉のT1値も含水量も減少したが、いずれも接種後10日から14日目にかけての減少が最も著しかった(図-1)。特に、T1は水の減少に先駆けて減少していることが明かになった。接種枝におけるT1と含水量の関係においても同様な傾向が認められた。対照区ではT1も含水量にも変化は認められなかった。ちなみに針葉の褐変が顕著に認められたのは接種後3週間目だったことから、T1の変化は非常に早かった。これまでの数多くの研究で報告された本病に対するマツのどの反応よりも早くあらわれており、水プロトンNMR緩和時間の測定によって得られるデータは、非破壊的診断法としてさらには発病機構解明のための重要なパラメータになりうると思われる。
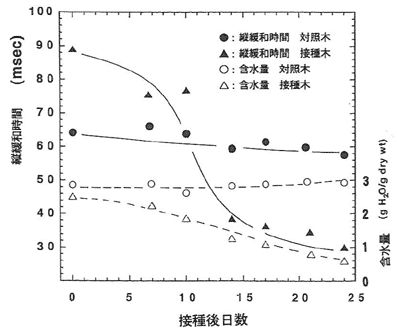
図-1マツノザイセンチュウ接種後のクロマツの針葉の縦緩和時間と含水量の変化
15.ヒノキ漏脂病発生林分の環境的特徴と罹病木における傷害の履歴
―京都府舞鶴市の4林分における調査結果―
黒田慶子(樹病研究室)
1. はじめに
ヒノキ漏脂病罹病木では、樹幹の二次師部内に傷害樹脂道が多層形成され、樹脂流出が継続し、形成層の部分的壊死による樹幹の変形が起る。発病原因究明のためには傷害樹脂道が繰り返し形成される条件を明らかにする必要がある。しかし漏脂症状が罹病後年月を経て出ることから、患部で腐生的な菌が繁殖するなど二次的な変化もあって、発病要因についての情報が得にくくなっている。被害林分の環境条件と罹病程度との関係について調べると同時に、罹病初期の状況を明らかにすることが重要と思われるので、枝打ちなど外傷の履歴と傷害樹脂道形成の関係について検討した。
2. 試料と方法
京都府舞鶴市の4箇所(17、22、24、32年生林分)で被害発生状況を調査した。また、漏脂症状と樹齢や枝打ち年度との関係、被害林分の標高、方位、傾斜について調べた。17年生1個体及び隣接する林分から14年生1個体、漏脂症状のある個体を伐倒した。さまざまな病患部から顕微鏡観察用切片を作製し、トルイジンブルー染色を行った。顕微鏡下で傷害樹脂道の形成程度を観察し、枝打ち年度と樹脂道形成開始年度との関連を明らかにした。さらに、漏脂患部付近から菌の分離を行い、漏脂症状と微生物感染との関わりについて検討した。
3. 結果と考察
土壌が乾燥気味の1林分では漏脂症状は見られなかったが、肥沃土壌の1林分では被害率80%に達した。他の2林分では斜面上部に被害がなく、谷下部の土壌が堆積している所に被害が集中していた。樹脂漏出程度は比較的軽微であったが、形成層の壊死がすでに12年前に開始していた個体もあった。伐倒した2個体は木部の年輪幅が5~8mmと成長が非常に良く、師部の年輪幅(図-1)も広かった。このように漏脂症状と肥沃地(成長過剰)との関連が示唆された。また、被害地の周囲では一部のスギに凍裂が見られ、著しい寒冷地ではなくても早春にかなりの気温の低下があるものと推測された。従って低温刺激と樹脂道形成との関連についても可能性は否定できない。14年生個体にはキバチ類の産卵があり、材の変色が認められた。産卵による傷害と樹脂道形成が対応するが、永続的な漏脂症状につながるかどうか明らかではない。
顕微鏡観察によると、17年生個体は樹幹全周に傷害樹脂道が形成されていた。樹皮内の傷害樹脂道は10年前の年輪および7~8年前の年輪から内側に向って各年輪に認められ(図-1)、特に枝打ち跡周辺に樹脂道形成が目立った。8~9年前の枝打ちによる傷との関連が示唆される。樹脂道の形成年輪から推定すると、罹病時期は樹齢8年ごろというかなり若い時期である。14年生個体では前年度形成の師部に樹脂道が認められ、木部内の傷害組織と対応した。前年の枝打ちおよびキバチ類産卵と樹脂道形成との関連が示唆されたが、キバチ類の産卵は漏脂病罹病後に起った可能性もある。伐倒木の漏脂患部周辺からは Penicillium, Cryptosporiopsis, Pestalotia, Sarea属菌及びキバチ類の産卵に伴って感染するAmylostereum属菌が検出された。

図-117年生罹病木横断面
右側の数字は師部の年輪(当年形成=0)を示す。
R: 樹脂分泌能力のある傷害樹脂道。
16.ヨゴオナガコマユバチにおける寄主サイズに応じた寄生数と性比の変化
浦野忠久・藤田和幸・上田明良(昆虫研究室)
1. はじめに
スギカミキリの寄生バチであるヨゴオナガコマユバチ(Doryctes yogoi Watanabe)は、丸太接種したスギカミキリ幼虫に対して高い寄生率を示すことが明らかにされている。本種の繁殖生態を明らかにするため、室内飼育を行い、寄主1頭あたりの寄生数及び次世代の性比が、寄主サイズとどのような関係にあるかを調査した。
2. 材料と方法
3月に野外で採集したスギカミキリ成虫を5月上旬にシャーレ内で産卵させ、20℃一定で保存して孵化幼虫を得た。6月初旬、スギ丸太(長さ80cm、直径10~15cm)に1本あたり20頭接種し、塩ビ円筒に1本ずつ入れてヨゴオナガコマユバチ雌成虫1頭を放ち、1週間産卵させた。異なるサイズの寄主幼虫に寄生させるため、産卵試験は接種10日後から50日後まで、10日おきに5回行った。接種丸太は1回の試験に2本ずつ、計10本を供試した。試験終了2週間後に供試木を剥皮し、樹皮下の寄生バチ次世代(多くは繭)と摂食後の寄主残骸(頭部)を取り出し、寄主残骸の頭幅を測って寄主サイズとした。次世代ハチは室内で羽化後雌雄を判別した。
3. 結果と考察
8本の供試丸太において3~8頭のスギカミキリ幼虫が寄生を受け(平均4.9頭)、残り2本では1頭も寄生されていなかった。雌1頭の産んだ次世代総数は3~18で、平均11.9であった。供試雌(丸太)ごとの寄主頭幅と寄主1頭あたりの寄生(繭)数の関係を図-1に示した。寄主サイズが大きくなるとともに寄生数の最大値が上昇し、それに伴ってばらつきも大きくなった。図には示していないが、雌ごとに両軸の値の平均をとると、右上がりの直線関係となった。寄主サイズが増加すると、1頭あたりの発育可能なハチ数が増えるものと考えられるため、雌バチは寄主サイズに応じた産卵数の調整を行ったとみることができる。寄主サイズとともに寄生数のばらつきが大きくなった原因としては、寄主摂食の段階で一部のハチ幼虫が死亡したか、あるいは雌バチの保有卵数があまり多くはなく、試験期間中に栄養摂取ができなかったために、新たな卵成熟がなされず産卵数が制限されたことが考えられる。供試雌ごとの寄主頭幅平均値と次世代性比の関係を図-2に示した。比較的大きな寄主を与えられた雌では、次世代の性比は極端に雌に偏っていた。一方寄主サイズが小さい場合は、産卵数と性比がトレードオフに近い関係にあるものと思われ、すべてが単寄生であったNo.1では、次世代3頭のうち2頭が雌であったのに対し、寄主1頭に2個体以上寄生する割合の高かったNo.2及び3では雄率が高かった。
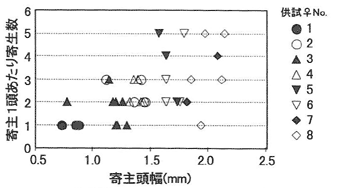
図-1 供試雌個体別に見た寄主サイズと寄主1頭あたり寄主(繭)数の関係
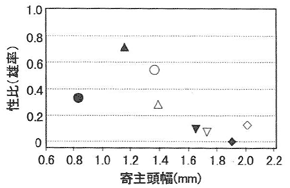
図-2 供試雌個体別に見た平均寄主サイズと次世代羽化成虫の成否の関係
(供試♀No.は図-1と同じ)
17.オオコクヌストの産卵選好性
上田明良・藤田和幸・浦野忠久(昆虫研究室)
1. はじめに
マツノマダラカミキリ(以下マダラ)の捕食性天敵オオコクヌスト(以下オオコク)の成虫は、マダラ同様マダラコールを用いたトラップで多数採集されることから、マツ類枯死木の臭いに誘引されると考えられる。しかし、本種のマツ類枯死木到着後の行動については知られていない。そこでオオコクの産卵行動へのマダラ幼虫の有無の影響を調べた。
2. 研究方法
1996年7月3日にアカマツを伐倒し、1mに玉切りして枝を切り落としたのち、オオコクの産卵を誘導するため、カッターで内樹皮へ至る傷を8箇所ずつつけた。この丸太30本のうち15本には7月8~22日のあいだ野外網室内でマダラに産卵をさせ、22日以後は常温の室内で保存した。残りの15本は伐倒日以後そのまま常温の室内で保存した。7月31日に上記2つの処理丸太の3本ずつ計6本を5つの野外網室へ分け入れ、交尾後のオオコク雌成虫を各網室に2頭ずつ放した。放虫から5日後の8月5日に丸太を取り出してオオコクの産卵状況を調べた。
3. 結果と考察
場所別にみた産卵箇所数と産卵数を表-1に示した。産卵箇所数と産卵数ともに折れまたは切り枝元の内樹皮下に多くみられた。折れまたは切り枝元の内樹皮は剥ぎにくく、オオコクにとっては安定した産卵場所と考えられる。これに対し外樹皮は剥がれやすく、産卵を敬遠した可能性がある。今回は根元近くの幹で樹皮の厚い部分の丸太を用いなかったので、こういった外樹皮の削ぎにくい場所での産卵については今後の検討が必要である。カッターでつけた傷の下にはまったく産卵がなかった。内樹皮に至る傷はオオコクの産卵には深すぎるのか、それとも樹脂を避けたのかは不明である。
マダラ産卵の有無別にみたオオコク産卵箇所数と産卵数を表-2に示した。全てのケージでマダラ産卵ありの丸太のほうがなしの丸太よりも産卵箇所数が多く、ケージ1を除けば産卵数も多かった。枝元あたりの産卵率(表中のB/A)と産卵数(表中のC/A)がいずれもマダラ産卵ありのほうが大きかったことから、オオコクはマダラ幼虫の存在する丸太をより選好したと考えられる。
これまでの結果と今回の結果を合わせると、オオコクは、まずマツ枯損木の臭いに定位し、つぎに丸太内の寄主の存在を確認して、最後に産卵に適した場所(樹皮薄い部分では折れ枝元の樹皮下)を探し産卵すると考えられる。
| 人為傷下 | 外樹皮下 | 枝元の樹皮下 | |
| 産卵箇所数 | 0 | 3 | 19 |
| 産卵数 | 0 | 32 | 84 |
| 平均卵塊サイズ | - | 10.7 | 4.4 |
| 網室No. | マダラ産卵 | 枝元数 | マダラ幼虫数 | 産卵箇所数 | 産卵数 | 枝元産卵箇所数 | 枝元産卵数 | B/A | C/A |
| (A) | (B) | (C) | (%) | ||||||
| 1 | あり | 39 | 24 | 4 | 20 | 4 | 20 | 10.3 | 0.51 |
| なし | 26 | 0 | 2 | 20 | 1 | 3 | 3.8 | 0.12 | |
| 2 | あり | 30 | 36 | 4 | 15 | 4 | 15 | 13.3 | 0.50 |
| なし | 24 | 0 | 1 | 9 | 1 | 9 | 4.2 | 0.38 | |
| 3 | あり | 34 | 22 | 1 | 10 | 1 | 10 | 2.9 | 0.29 |
| なし | 35 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 4 | あり | 47 | 35 | 2 | 8 | 2 | 8 | 4.3 | 0.17 |
| なし | 23 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 5 | あり | 29 | 30 | 8 | 34 | 6 | 19 | 20.7 | 0.66 |
| なし | 19 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
18.マツノマダラカミキリの捕食性昆虫による密度抑制効果
上田明良・藤田和幸・浦野忠久(昆虫研究室)
1. はじめに
マツノマダラカミキリ(以下マダラ)幼虫の天敵として寄生蜂や捕食性昆虫及びキツツキ類が知られている。マダラの防除のためには、これら天敵の誘致・定着、あるいは放飼が期待される。その前に現実に果たしている役割を把握することは重要である。しかし、寄生蜂や捕食性昆虫のマダラ幼虫に対する密度抑制効果がどれほどかを野外で実験的に調べた例はない。そこで、今回捕食性昆虫による密度抑制効果をみるための試験を行った。
2. 研究方法
1995年7月6日に伐採したアカマツを約1mに計40本玉切りし、野外網室内で7月28日までマダラに産卵させたのち。滋賀県野洲町のアカマツ山林内4箇所に10本ずつ設置した。各区10本のうち5本は1本ずつ二重の寒冷紗製袋に入れて天敵の侵入を防ぎ、裸の丸太と交互に立て掛けた。同年12月7日に丸太を割材し、マダラ蛹室数と個体数を調べた。
3. 結果と考察
結果は表-1に示すように、尾根付近のアカマツ林に設置した場所B、C、Dでは蛹室数の合計が寒冷紗袋に入れた方と入れなかった方とでほぼ同数(場所B)、またはむしろ入れなかった方が多かった(場所CとD)ことから、マダラ幼虫が蛹室を作るために材入するまでの樹皮下での生活中に寄生や捕食をほとんど受けなかったと考えられる。これに対し、マダラの空室の蛹室率については、場所B、C、Dともに寒冷紗袋に入れなかった方が高く、また場所BとDでは空室数に寒冷紗袋に入れた方と有意差があったことから、マダラ蛹室内幼虫への捕食効果があったと考えられる。場所BとDではオオコクヌスト(以下オオコク)幼虫がマダラ蛹室から採集された。丸太にはキツツキ類が辺材をつついた痕跡はなく、空室内にいたであろうマダラ幼虫のほとんどはオオコクに捕食されたものと考えられる。寒冷紗のある区とない区の空室率の差が被食率と仮定すると、場所Bで63.5%、Cで34.4%、Dで59.0%の蛹室内幼虫が捕食されたことになる。
山麓の場所Aでは空室のマダラ蛹室率には違いがなかったことから、場所B~Dのような蛹室内のマダラ幼虫への捕食はなかったと考えられる。しかし、他の場所と違って、寒冷紗袋に入れた方が有意にマダラ蛹室数の合計が多かった。
| 設置場所の環境 | 天敵侵入の阻止 | マダラ数(5本からの合計) | B / (A+B) | ||||
| 樹皮下幼虫 | 材内幼虫 | 空室の蛹室 | 蛹室計 | (%) | |||
| (A) | (B) | (A+B) | |||||
| A | 山麓の密なアカマツ林 | あり | 0 | 32* | 3 | 35* | 8.6 |
| なし | 3 | 8 | 1 | 9 | 11.1 | ||
| B | 尾根近くの密なアカマツ林 | あり | 0 | 30* | 1** | 31 | 3.2 |
| なし | 1 | 9 | 18 | 27 | 66.7 | ||
| C | 尾根近くの粗なアカマツ林 | あり | 1 | 17 | 1 | 18 | 5.6 |
| なし | 2 | 18 | 12 | 30 | 40.0 | ||
| D | 尾根近くの粗なアカマツ林 | あり | 0 | 21 | 0* | 21 | 0 |
| なし | 6 | 16 | 23 | 39 | 59.0 | ||
*天敵にさらした方とU検定で5%水準の有意差あり。**同1%水準。
このことはこの場所では樹皮下での生活中に寄生や捕食を多く受けたことを示唆する。寄生蜂の繭がひとつもみつからなかったことから、場所Aは樹皮下幼虫への捕食者が作用しやすい環境であったのかもしれない。今後の検討が必要である。
場所B~Dのような尾根近くでは毎年数多くのマツ枯れがみられたのに対し、場所Aのある山麓での枯れはほとんどなかった。オオコク成虫はマダラコールを用いたトラップに多数誘引されることから、マツ枯損木が多いところにより強く誘引され、このことが尾根近くと山麓の間で、マダラ蛹室内幼虫に対するオオコクによる捕食効果の違いをもたらしたのかもしれない。
19.京都府下におけるツキノワグマ駆除個体群の年齢構成
北原英治(鳥獣研究室)
1. はじめに
我が国においてツキノワグマ(Selenarctos thibetanus)は本州と四国に生息し、西日本では深刻な樹木の剥皮被害を引き起こすことから地域によっては有害駆除が実施されている。しかし、一方では様々な人為インパクトにより個体群の分断化が起こり、西日本のツキノワグマはその存続が危ぶまれている。京都府下でも地域個体群の絶滅が危惧されながらも、被害軽減の目的から長年駆除が実施されている。これらの駆除個体の解析のうち、本年度は個体群の年齢構成について調査を行った。
2. 方法
京都府各地方振興局から当研究室へ送付された冷凍試料(1991~1995年、49頭)から、ツキノワグマの上顎犬歯を抜歯して年齢査定のための試料とした。抜歯された犬歯は縦方向に短冊型に歯科用のデイスクで切断して、切片作成のための試料とした。なお、脱灰はプランクリチュロ液で行い、検鏡用の試料は厚さ30~40μmで作成し、染色はカラッチヘマトキシリン液にて行った。
3. 結果と考察
調査の結果、京都府下のツキノワグマ駆除では当歳から22歳までの個体が駆除されていた(図-1)。まず、歯根部分がほとんど伸長していない、歯冠部分のみの犬歯を有する当歳個体は、1992年に親子連れ個体として捕獲されていた。また、年越しの1歳個体は、1991年と1995年に単独で駆除されており、やはり歯根が開放した犬歯を有していた。最終的に歯根部分が完全に閉鎖するのは5~6歳であることが分かった。
次に、個体群の年齢構成として見ると3歳から7歳までの個体が全体の約65%を占め、京都府では盛んに駆除の対象となっていた。最高齢22歳個体(雄)は兵庫との府県境に位置する久美浜町で1992年に、また16歳個体(雌)は夜久野町で1991年に駆除されていた。さらに、13歳個体(雄)は福知山市で1991年に、12歳個体は雌が美山町で1992年に、雄が久美浜町で1994年にそれぞれ駆除されていた。京都府下で駆除されるツキノワグマ個体群は宮津・舞鶴群と美山・京北群の2群が認められているが、年齢構成を概観すると上記の通り宮津・舞鶴群でより老齢な個体が駆除される傾向が認められた。なお、経年における駆除個体の若齢化などの変化が今回の分析からは明らかにされなかった。
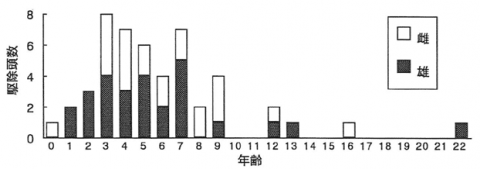
図-1 京都府下における駆除個体の年齢構成
20.ヒメネズミによるブナ堅果の分散貯蔵
島田卓哉(鳥獣研究室)
1. はじめに
堅果類の散布は、もっぱら動物による「貯食型散布」によって行われている。貯食型散布は分散貯蔵された堅果が、動物に回収され忘れて初めて実現する。したがって、堅果類の動物による散布の効果を評価するためには、分散貯蔵される割合、分散貯蔵される場所(発芽・成長の適地か)、回収される割合などを知ることが重要である。筆者は、奈良県大台ヶ原においてブナ堅果とヒメネズミを対象として調査を行っている。本稿では、その中でも堅果が分散貯蔵される場所の距離、方向、条件について報告する。
2. 方法
調査は、奈良県大台ヶ原の針広混交林で行った。高木層には、ブナ、ミズナラ、ウラジロモミが優占している。調査地域内にはアカネズミとヒメネズミが生息しているが、調査を行った11月上旬にはヒメネズミのみが捕獲された。
一端に標識テープを結んだ針金を健全なブナ堅果に取りつけた。これをブナ林冠下の地表に30個置き、その上に鳥が入れないようにプラスチックのカバーを施した。このようなサイトは2箇所に設けた。1回目の実験では、針金付堅果を1996年11月7日に設置し11月8日に利用状況をチェックした。2回目の実験は、11月8日設置、11月13日チェックである。いずれの実験でも、設置場所から分散貯蔵された場所までの距離と方向、どのような場所であったかを記録した。
3. 結果と考察
30個の堅果の内、持ち去られた堅果の個数は、サイト1では27(11月7日設置分)及び26(11月8日設置分)、サイト2では3(11月7日設置分)及び5(11月8日設置分)であった。その内分散貯蔵が確認されたのは、サイト1では10(11月7日設置分)及び8(11月8日設置分)、サイト2では3(11月7日設置分)及び1(11月8日設置分)であった。ネズミの貯食行動については、体重や齢がその行動に影響することが報告されている。たとえば、Nakatsuyama & Fujita(1995)は、ラットについて雌の方が雄よりも餌を運搬する傾向が強いことを報告している。サイト1と2で持ち去られた堅果数に大きな違いが認められたのは、ヒメネズミの条件が制御されていなかったことに原因がある可能性がある。
散布された堅果の距離と方向を図-1に示した。いずれのサイトにおいても、分散貯蔵はある一定の方向に集中してなされている。今回の調査ではネズミの行動圏を評価するのに十分なデータは得られていないが、おそらくここで見られた分散の定向性はヒメネズミの活動中心と関連しているものと思われる。分散された距離は、たかだか7m程度であった。より遠くに分散された堅果を見落としている可能性は否定できないが、分散されても樹冠内に留まる場合も多いことが示された。
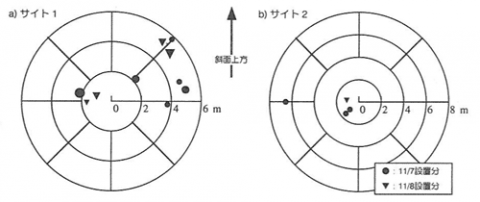
図-1 分散貯蔵された堅果の距離および方向
記号が分散貯蔵が確認された場所. 記号の大きさは貯蔵されていた堅果数を示している.
分散貯蔵された堅果22粒の内、19粒が倒木下、根株下、地表に露出した根の下などに持ち込まれていた。堅果類は乾燥に弱く、地表に置かれていると1週間程度で発芽能力を失ってしまう(斎藤、1992)。そのため、堅果が発芽に成功するためには、ネズミなどによって乾燥しにくい場所に分散されることが重要である。本研究では、ヒメネズミによる分散貯蔵においては、堅果の多くは倒木下などの発芽のための適地に散布されることが示された。
引用文献
- Nakatsuyama, E. and Fujita, O. (1995) J. Ethology,13, 95-103.
- 斎藤新一郎 (1992) 生物科学,44(2), 89-97.
21.ツキノワグマによる林木剥皮被害
北原英治(鳥獣研究室)
井上重紀・今井三千穂(福井総合グリーンセ)
山田文雄(森林総研森林生物部)
島田卓哉(鳥獣研究室)
小泉 透(森林総研九州)
1. はじめに
従来、ツキノワグマによる林木の剥皮被害(クマ剥ぎ)は西日本の太平洋側の地域に多く見られ、西日本で日本海側や東北地方には極めて少ないと言われていた。しかし、最近の調査によって西日本の日本海側や東北地方においてもクマ剥ぎ被害は深刻な実態にあることが明らかにされた(大津・大泉、1991)。一方、ツキノワグマはその生息環境の改変などから個体数の急激な減少が心配され、特に人工林化の進んだ西日本では地域的な絶滅も案じられている。林業との共存の意味からも被害防除技術の確立が強く要望されている。そこで、先ず有害駆除による個体数調整が行われている地域での被害実態を明らかにして、被害発生の要因について考察する。
2. 調査地の概況と調査の方法
福井県名田庄村の森林所有者を対象に聞き取り調査から、クマ剥ぎの激害地域を選定し、そこに調査地(20~27年生)を設定した。名田庄村は人工林率が高く、いわゆる天然林が尾根筋に残っているのみである。スギ造林地は地形的に斜面上部から尾根部、斜面中部及び斜面下部に類型されるが、調査プロットA~Eはそれらを含む形で設定された。プロットの大きさは調査木の本数で決定され、プロットあたりの本数を30~35本とした。調査は1990年12月と1994年5月に実施され、剥皮後の樹皮の巻き込み状況から剥皮発生年を特定した。さらに、この地域で有害駆除によって捕獲されたツキノワグマ個体数を調べた。また、調査地において1994年5月に新鮮な糞を収拾し、その内容物を分析した。
3. 結果及び考察
1). 剥皮被害と有害駆除の実態
プロット毎の剥皮割合は、Aで93%(28/30本)、Bで90%(27/30本)、Cで77%(27/35本)、Dで80%(24/30本)、Eで33%(10/30本)であった。被害は中・大径木主幹における樹皮剥ぎの形態で発生し、剥皮被害木の木質部には切歯(または犬歯)による柔らかい形成層部分の削り落とし跡が見られた。剥皮部分に生じた樹皮の巻き込み状況から推定した被害発生年をプロット毎に示した(図-1)。調査実施が1990年と1994年であったため、年区分はa;1~2年前、b;3~5年前、c;6~10年前、d;11~15年前及びe;16年以上前、とした。それらによると、剥皮被害は、A、B、C及びDの激害地ではほぼ全年区分で発生していた。次に、この地域における有害駆除によって捕獲されたツキノワグマの個体数について見ると(図-2)、1989年の10頭を最高に、毎年捕獲されていた。捕獲個体の性比を見ると、1987年と1990年(1991年と1992年は性別不明)を除くと、いずれの年も雄個体が多く捕獲されており、剥皮被害の発生する時期がかれらの交尾期に当たっていて雄個体が活発に行動する(ブロムレイ、1972)結果、捕獲されたものと思われた。このように、毎年多数の個体(特に雄)が捕獲されているにもかかわらず、剥皮被害の発生がおさまる傾向がみられないことから、有害駆除及び一般狩猟による個体数調整は必ずしも被害の軽減化に関与していないように思われた。実際、最近のテレメトリーなどによって調べられたツキノワグマの行動圏は雌で1.2~2.5km2に対して雄で60km2とされ(石田ら、1993)、季節的な移動を考慮すれば、1地域における被害の軽減を目的とした有害駆除効果には限界があると考えざるを得ない。さらに、被害防止法としての集中的な駆除は地域的な個体群の孤立化や絶滅を助長させることになると思われた。
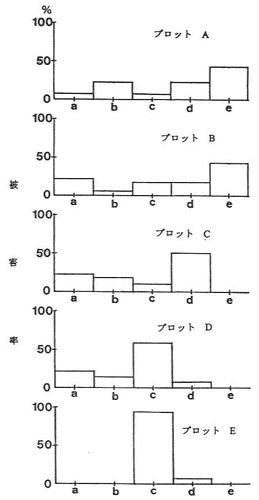
図-1 剥皮被害の発生年別頻度
a; 1~2年前, b; 3~5年前,
c; 6~10年前, d; 11~15年前,
e; 16年以上前
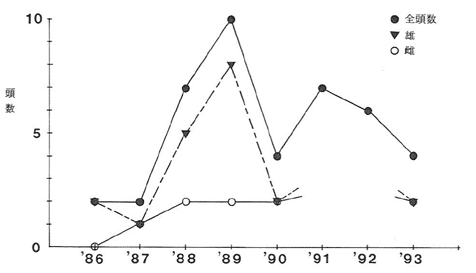
図-2 名田庄村におけるクマの捕獲個体数の推移
2). 糞分析の結果
剥皮被害の発生地にて5月に収拾したツキノワグマの糞を分析した結果、識別不能な緑小片に紛れて木質小片(形成層)が多数検出された。樹木の剥皮被害発生時期は図-3に示した食物カレンダーに組み込まれており(水野・野崎、1985)、今回、この被害が樹木の粗皮下にある柔らかい形成層部分をそぎ落として摂食するために起こることが判明した。水野・野崎 (1985)によれば、この時期、クマは主に林内に豊富にある樹木の花、芽及び葉などを大量に摂食しており(図-3)、絶対的な食物不足から樹皮剥ぎを起こすのではなく、好んで摂食していることが推測された。
1)の結果から、ツキノワグマにおいてもニホンジカやニホンザルなどの大型獣類と同じく、その個体群に対するモニタリングと、それに基づく総合的な保護管理体系の確立が重要であり、捕殺によらないソフトな被害防止法の確立が必要であることがわかる。具体的には、各地でその効果が試験されているテープ巻き、臭覚を刺激して忌避させる化学的方法や、樹皮剥ぎの発生しない生息環境への環境の改変(例えば、2)の結果から考えられる1つの方法として、スギよりも嗜好性の高い樹種があればそれを植林し、スギへの被害を減らす)などについてのデータの集積が必要であろう。
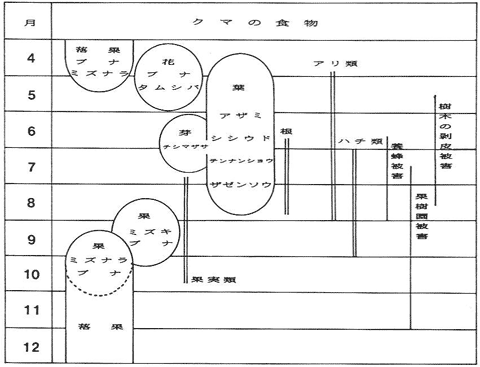
図-3 白山山系におけるクマの食物(丸囲いは重要な食物)
(水野・野崎、1985を改変)
引用文献
- ブロムレイ、G. F.(1972) 南部シベリアのヒグマとツキノワグマ,(藤巻裕蔵・新妻昭夫、訳)、北苑社、札幌.
- 石田 健・赤石朋敏・山中隆平(1993) 東京大学秩父演習林における道路開設が野生鳥獣の生態に与える影響(III)-ツキノワグマの捕獲・標識・行動圏調査-, 104回日林論, 695~696.
- 水野昭憲・野崎英吉(1985) 白山山系のツキノワグマの食性,「森林環境の変化と大型野生動物の生息動態に関する基礎的研究(環境庁自然保護局)」, 38~43.
- 大津正英・大泉雅春(1991) ツキノワグマによる林木等の被害,日林東北支論、43, 157~158.
Copyright © Forest Research and Management Organization. All rights reserved.