
研究紹介 > トピックス > プレスリリース > プレスリリース 2022年 > 奥尻島のブナ集団は最終氷期最盛期以前に形成された ~ブナの北方への分布移動の歴史~
ここから本文です。
![]()
2022年12月15日
国立大学法人筑波大学
国立研究開発法人森林研究・整備機構 森林総合研究所
十日町市立里山科学館 越後松之山「森の学校」キョロロ
国立大学法人北海道教育大学
奥尻島のブナ集団は最終氷期最盛期以前に形成された
~ブナの北方への分布移動の歴史~
ブナは日本を代表する冷温帯樹種で、南は鹿児島県大隅半島の高隅山から北は北海道黒松内周辺まで分布しています。本研究グループではこれまでに、道南~黒松内および以北に隔離分布しているブナの分布最前線と、これら集団の遺伝的多様性や集団形成の歴史を明らかにしてきました。
本研究では、まだ解析されていなかった、北海道奥尻島のブナ集団に焦点をあて、東北地方~北海道本島も含めて、母性遺伝する葉緑体DNAおよび両性遺伝する核DNAを用いて、集団遺伝学的解析を行いました。その結果、奥尻島には全国で見られる日本海側系統と太平洋側系統の両系統のブナが分布していました。集団動態の歴史の推定からも、奥尻島のブナ集団は、北海道および東北地方集団双方からの混合により形成され、その後も北海道、東北地方から遺伝的交流があったことが示唆されました。また、奥尻島の最も古いブナ集団は、最終氷期最盛期(約2万年前)より前に形成された可能性が高いことが分かりました。これは、花粉解析など古生態学的な先行研究や奥尻島の地史を支持しています。さらに、現在は離島として隔離されていますが、その遺伝的多様性は北海道、東北地方の集団と同程度でした。これらのことから、奥尻島のブナ集団は、複数回にわたる個体または種子、花粉の移動や、他地域からの遺伝的交流によって形成されたことが明らかになりました。
本研究成果は、ブナが北限地域へどのように分布したかを理解する上で重要であるだけでなく、今後の冷温帯林の気候変動影響評価にも資するものです。
研究の背景
離島に生育する植物集団は、地理的に隔離されているために、分布の移動や拡大、生残などの生態学あるいは集団動態的なプロセスが、地理的に連続している本島集団とは異なります。集団遺伝学的な観点からは、離島では、本島からの移動分散により集団形成される際に、創始者効果注1)やびん首効果注2)により、遺伝的多様性が低下すると考えられ、この仮説にあてはまる先行研究例は多くあります。また、離島集団は本島の連続分布集団に比べて分布拡大ルートが限られるため、過去の分布拡大の歴史が異なります。一方、離島集団でも本島集団と同程度の遺伝的多様性を示す報告もあります。これは本島の母集団からの遺伝子流動(花粉あるいは種子の流動)や環境選択など複数の要因があると考えられています。
ブナ(Fagus crenata)は日本を代表する冷温帯樹種で、南は鹿児島県大隅半島の高隅山から北は北海道黒松内周辺まで分布しています。先行研究からは、分布域の南から北へと遺伝的多様性が低下する地理的パターンがあること、核および葉緑体DNA、葉の形態に、太平洋側系統と日本海側系統があること、などが知られています。また本研究グループではこれまでに、道南~黒松内とそれ以北に隔離分布注3)しているブナの分布最前線、および、これら集団の遺伝的多様性や集団形成の歴史を明らかにしてきました(Kitamura et al. 2015、北村ら2016)。しかしながら、道南の離島である奥尻島は、調査対象に含まれていませんでした。
奥尻島は、本島から約16km離れ、北緯約42度に位置しており、ブナ集団の分布最北地域といえます(図1、2)。地質学的研究からは、奥尻島は中期更新世に北海道本島から分かれて以降、海面が現在よりも下がった最終氷期最盛期(約2万年前)を含めて、一度も本島とは陸続きになっていないと考えられています。花粉解析などを用いた古生態学的研究から、ブナの北進は最終氷期最盛期の後、今から約6000年前に始まったと考えられていますが、その母集団の由来なども分かっていませんでした。
そこで本研究では、奥尻島のブナ集団に焦点をあて、奥尻島および東北地方~北海道本島のブナ集団を対象に、集団遺伝学的解析注4)により、奥尻島のブナの集団動態と、最終氷期最盛期を含めた、ブナの北方への分布移動・拡大の歴史の解明に取り組みました。

図1 奥尻島のブナ林の様子(撮影:北村系子)
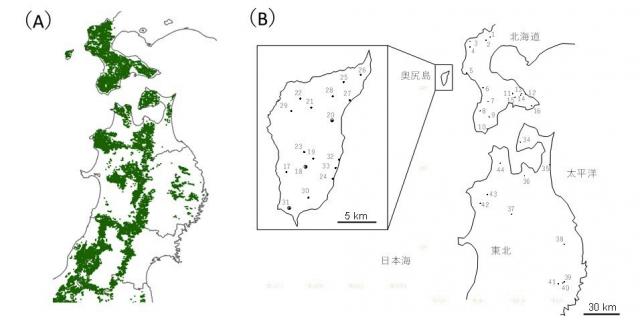
図2 (A)東北地方~北海道のブナの分布(Matsui et al. 2004を改変)および(B)本研究で用いたブナ集団の位置
研究内容と成果
本研究では、奥尻島の17集団、北海道の16集団、東北地方の11集団の計44のブナ集団より1838個体のブナの葉を採取し、遺伝様式の違いからより複雑な集団の歴史を推定できるように、母性遺伝する葉緑体DNAと両性遺伝する核DNAの2つの遺伝的変異情報を用いて、大規模な集団遺伝学的解析を行いました。その結果、葉緑体DNAからは、全国のブナに見られる日本海側系統(ハプロタイプ注5)A:図3赤色部分)と太平洋側系統(ハプロタイプB:図3黄色部分)の両系統が、奥尻島より検出されました。本研究グループの奥尻島集団を含まない先行研究から、北海道内では道南から分布最北の最前線に向けて遺伝的多様性が低下することが明らかになっており、これは最前線集団が比較的最近形成されたことによると考えられます。一方、本研究では、核DNAの遺伝的多様性は奥尻島、北海道、東北地方の3地域で同程度であることが分かり、奥尻島のブナ集団は、最前線集団形成よりも以前から集団を維持してきたことが示唆されました。
また、核DNAのデータから、全個体を2つのクラスター(遺伝的グループ)に分けると、大きくは東北地方を主とするクラスター(図4A:赤色部分)と北海道を主とするクラスター(図4A:黄色部分)の分布に地理的パターンがあり、奥尻島では両クラスターの混合注6)パターンが見られました。さらに全個体を3つのクラスターに分けると、奥尻島を主とする青色クラスターが検出されましたが、奥尻島から北海道と東北地方の両クラスターも見られるというパターンは、2クラスターに分けたときと同様の結果でした(図4B)。これらの遺伝構造を詳細に評価するために集団動態の歴史を推定したところ、奥尻島のブナ集団は、北海道集団と東北地方集団の混合により形成され、その後も北海道、東北地方のブナ集団からの遺伝的交流があったことが分かりました(図5)。特に興味深いことに、奥尻島の初期ブナ集団の形成は、最終氷期最盛期(約2万年前)よりも以前まで遡る可能性が高いことが示されました。これらの結果から、奥尻島のブナ集団は、かつて1度のイベントではなく、複数回に渡る移動イベントや他地域からの遺伝的交流により、現在の集団が形成されたと考えることができます。この知見は、花粉解析など古生態学あるいは地質学的な奥尻島に関する先行研究を支持し、奥尻島のブナ集団は最終氷期最盛期以前から奥尻島に分布していたことが、遺伝データからも明らかになりました。
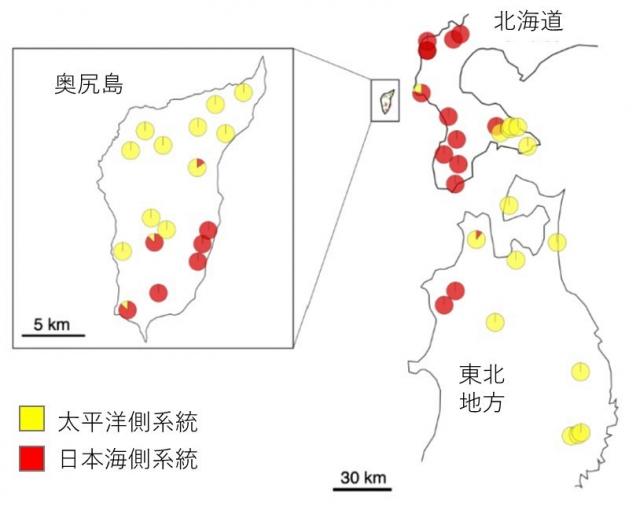
図3 母性遺伝する葉緑体DNAでみた2つのハプロタイプ(黄色・赤色)の分布
先行研究のFujii et al. (2002)によると、黄色ハプロタイプはブナの太平洋側系統に、赤色ハプロタイプは日本海側系統に対応する。本研究では両系統が日本海側の奥尻島から検出された。
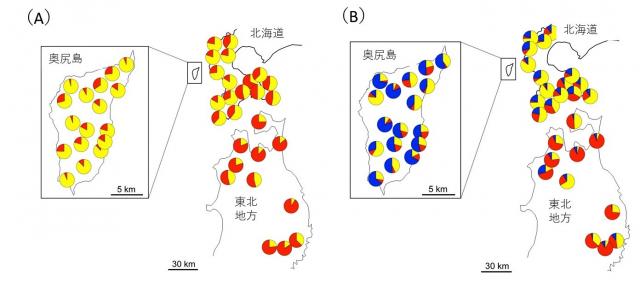
図4 集団内の核DNAに基づく遺伝的クラスターの分布
(A)2つの遺伝的クラスターに分けると、東北地方を主とするクラスター(赤色)と北海道地方を主とするクラスター(黄色)があり、奥尻島集団では混合パターンがみられた。
(B)3つの遺伝的クラスターに分けた場合、赤色、黄色、青色で示したクラスターはそれぞれ、東北地方、北海道、奥尻島に対応する。
いずれも複数クラスターの混合構造が見られるが、これは氷期ー間氷期における、ブナの分布移動に関連した集団動態の歴史や、その後の花粉、種子を介した集団間の遺伝的交流によるものと考えられる。
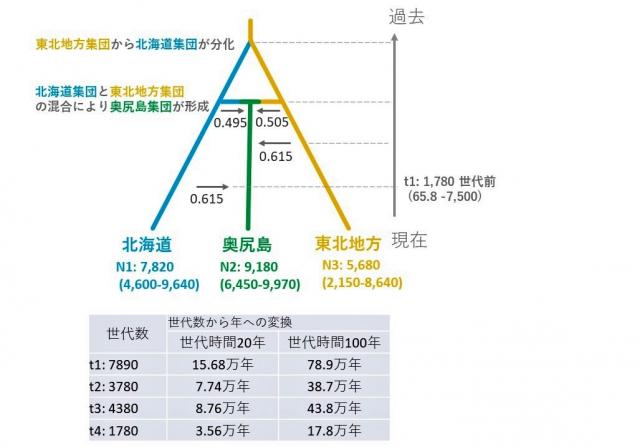
図5 推定された集団動態の歴史のモデル図
各集団のN数は有効集団サイズを、各推定値のカッコ内の数値は推定値の95%信頼区間相当(=統計的に意味のある値)の値を示す。図内の矢印は集団間の方向性のある混合を表す。図下部の表は、世代時間として20年および100年を仮定した場合の年変換。(時間は「世代数」で計算されるため、「年」への変換は仮定する世代時間に依存する。)
今後の展開
本研究結果は、ブナが北海道などの北限地域へどのように分布を拡大したかを理解する上で重要であるだけでなく、今後の冷温帯林の気候変動影響評価の上でも大きな成果と言えます。今後さらに、これらの地域のブナ集団の遺伝構造、集団動態、環境適応を、ゲノムレベルでより詳細に調べるとともに、ブナ属樹種の進化を全球レベルで評価すべく、海外グループとの研究を展開する予定です。
用語解説
注1)創始者効果
ある母集団から派生した隔離集団が形成される場合、限られた個体数から集団が形成されることにより、隔離集団が母集団とは異なる遺伝子の頻度を持つこと。一般に、豊富な遺伝的多様性をもつ母集団の中から限られた個体数を創始者として隔離集団が形成された場合、隔離集団の遺伝的多様性は母集団よりも低下していることが多い。(元に戻る)
注2)びん首効果
集団の個体数が大きく減少することで、個体数の限られた母集団から後代集団が形成され、集団の遺伝的多様性が低下すること。ボトルネック効果とも呼ばれる。(元に戻る)
注3)隔離分布
かつてはより広域に連続的に分布していた種が、過去の地殻、地形、気候などの変動により、分布が局所的になり、不連続になること。例えば、日本の高山植物はかつて現在よりも気温が低かった時期にはより広域に分布していたが、現在では高標高域にのみ隔離分布している。(元に戻る)
注4)集団遺伝学的解析
生物の集団内の個体間の遺伝的変異あるいは集団間の変異遺伝的などから、種~集団~個体レベルで遺伝的多様性を評価し、また集団動態の歴史などを推定する解析。(元に戻る)
注5)ハプロタイプ
ブナは2倍体生物であり、核ゲノムは2組の相同なゲノム組成を持つ。一方、全植物で葉緑体ゲノムは半数体(ハプロイド)であり、半数体を扱う場合の遺伝子型(ジェノタイプ)をハプロタイプと呼ぶ。(元に戻る)
注6)混合
異なる遺伝的組成(例えば北海道系統と東北地方系統)を持つ2個体が交配することで、その子供では、親の遺伝情報が半分ずつ混ざり合ったゲノム組成になること。実際の生物集団では、混合個体同士の世代を超えた交配や、混合が進んだ個体と親の純系統由来個体との交配(戻し交配)が繰り返されるため、同じ種であっても個体レベルでは種内系統の混合が見られる。(元に戻る)
研究資金
本研究は、日本学術振興会(JSPS)・科研費基盤研究C(JP17K07852, JP20K0615)、JSPS研究拠点形成事業B. アジア・アフリカ学術基盤形成型(JPJSCCB2022000)、公益財団法人自然保護助成基金プロ・ナトゥーラ・ファンド助成 、研究拠点形成事業アジア・アフリカ学術基盤形成型の一環として実施されました。
掲載論文
論文名:Possible northern persistence of Siebold’s beech, Fagus crenata, at its northernmost distribution limit on an island in Japan Sea: Okushiri Island, Hokkaido(分布北限の日本海・奥尻島のブナ集団の最終氷期最盛期の北方生残の可能性)
著者名:Keiko Kitamura#(国立研究開発法人森林研究・整備機構森林総合研究所・北海道支所)、Kanji Namikawa(北海道教育大学教育学部札幌校)、Yoshiaki Tsuda#(筑波大学生命環境系/山岳科学センター菅平高原実験所)、Makoto Kobayashi(十日町市立里山科学館 越後松之山「森の学校」キョロロ)、Tetsuya Matsui(国立研究開発法人森林研究・整備機構森林総合研究所/筑波大学生命環境系)
#本研究に対し同等の貢献をした。
雑誌名:Frontiers in Plant Science
掲載日:2022年12月15日
DOI:10.3389/fpls.2022.990927
| お問い合わせ先 |
|
研究担当者: 広報担当者: |
関連資料
![]()
PDF形式のファイルをご覧いただく場合には、Adobe Acrobat Readerが必要です。Adobe Acrobat Readerをお持ちでない方は、バナーのリンク先から無料ダウンロードしてください。
Copyright © Forest Research and Management Organization. All rights reserved.